转录因子E2F4对哮喘相关基因Six1的转录调控
2021-07-28周国平
刘 爽,陈 曦,周国平
南京医科大学第一附属医院儿科,江苏 南京 210029
支气管哮喘(简称哮喘)是儿童最常见的慢性炎症性疾病之一[1],在世界范围内的患病率呈上升趋势[2]。哮喘的发病机制复杂,包括免疫因素、感染因素、环境因素和遗传因素[3]。支气管高反应性、间歇性气流受限和气道重塑是哮喘的3 大典型特征。气道重塑是指长期反复的炎症不断刺激,诱发气道组织增生而出现反复修复,最终导致气道结构的改变,以气道壁增厚、上皮下纤维化、平滑肌数量增加、血管生成和黏液腺的增加为其主要特征[4]。当前,临床上尚无有效方法治愈哮喘。因此,重要的是探索气道炎症和重塑的机制,并寻求新的策略。
同源异型盒基因1(sine oculis homeobox homolog 1,Six1)是一种包含同源结构域的转录因子,对细胞增殖、迁移、侵袭和上皮-间充质转化(epithelial-mesenchymal transition,EMT)起着重要作用[5-7]。Six1表达在胚胎发生后大幅度下调,但在许多癌症中重新高表达,从而有助于肿瘤的生长[8-9]。在哮喘中,Six1对转化生长因子β(transforming growth factor-β,TGF-β)1 介导的EMT、气道炎症及气道重塑具有重要作用[10]。气道炎症和重塑是哮喘两个最典型的病理特征,渗透到气道的炎症细胞通过促进气道平滑肌细胞的增殖和迁移而有助于气道重塑。
因此,研究人Six1基因对哮喘、肿瘤等疾病有重要的理论指导意义。但是目前对Six1的研究主要集中在信号通路及其与蛋白的相互作用上,对于Six1自身的转录调控机制研究较少。本实验通过构建人Six1 启动子荧光素酶报告重组质粒,通过瞬时转染HEK-293细胞和BEAS-2B细胞,分析其在这2种细胞中的荧光素酶活性,接着通过qRT-PCR和蛋白质免疫印迹实验测定敲低、过表达E2F转录因子4(E2F transcription factor 4,E2F4)后Six1表达量的改变,旨在为进一步研究人Six1基因的转录调控机制奠定基础。
1 材料和方法
1.1 材料
人胚肾细胞(HEK-293)、人肺支气管上皮细胞(BEAS-2B)、pGL3-basic 质粒、pRL-TK(海肾荧光素酶报告质粒)为本实验室保存;E2F4 小干扰RNA siE2F4 及其阴性对照siNC、E2F4 过表达质粒pENTER-E2F4及空载体pENTER(南京擎科生物科技有限公司);DMEM 培养基(ThermoScientific 公司,美国);胎牛血清(Gibco公司,美国);青链霉素双抗(上海碧云天生物技术有限公司);小量质粒抽提试剂盒(Omega公司,美国);DNA Marker、限制性内切酶MIUⅠ和XhoⅠ、T4 DNA 连接酶和逆转录酶(TaKaRa 公司,日本);SYBR Green qPCR Master Mix(Bimake 公司,美国)。质粒转染试剂Lipofectamine3000脂质体(Invitrogen公司,美国);双荧光素酶检测试剂盒(Promega公司,美国);琼脂糖(GENE公司,西班牙);总蛋白提取试剂盒(南京凯基生物技术股份有限公司);E2F4、Six1、GAPDH 一抗(Proteintech 公司,美国);SDS-PAG 凝胶、辣根过氧化物酶标记羊抗兔二抗(上海碧云天生物技术有限公司)。
1.2 方法
1.2.1 细胞培养
HEK-293 和BEAS-2B 细胞在含有10%胎牛血清的DMEM 高糖培养基中培养,并置于37 ℃、5%CO2培养箱,2~3 d 传代1 次。实验操作均采用对数期生长细胞。
1.2.2 人Six1 启动子荧光素酶报告基因重组质粒的构建与鉴定
人Six1 基因序列(GenBank 编号:NC-000014.9)从美国国立生物技术中心数据库NCBI(http://www.ncbi.nlm.nih.gov/gene/)找到起始密码子ATG 上游约351 bp 片段序列。将人Six1 启动子的转录起始位点(TSS)设置为+1。将人Six1 DNA片段-351~100 nt的启动子插入pGL3-Basic 载体中,命名为pSix1-451。用JASPAR 软件(http://jaspar.genereg.net/)预测转录因子结合位点。
1.2.3 脂质体转染细胞
将HEK-293 和BEAS-2B 细胞分别接种到96 孔板中(1×104个/孔)。24 h 后,使用Lipofectamine 3000 将100 ng 启动子报告质粒pSix1-451 或pGL3-Basic 与pRL-TK 质粒(4 ng)一起共转染到细胞中作为内部对照。使用Lipofectamine 3000试剂将100 ng启动子报告质粒pSix1-451 与siE2F4(50 nmol/L)或siNC(50 nmol/L)瞬时转染到细胞中,于37 ℃、5%CO2培养箱培养24 h,观察敲低E2F4对pSix1-451启动活性的影响;另外,将pSix1-451与pENTER-E2F4(100 ng)或pENTER(100 ng)进行共转染,观察过表达E2F4对pSix1-451启动活性的影响。
1.2.4 双荧光素酶报告基因系统检测启动子活性
弃培养基,用100 μL/孔磷酸盐缓冲液(phosphate buffer saline,PBS)将96 孔板清洗2~3 遍,每孔加入30 μL 裂解液后室温震荡20 min,按照双荧光素酶检测试剂盒步骤测定pGL3-basic、pSix1-451以及敲低和过表达E2F4后pSix1-451的启动子活性,并将其标准化为pRL-TK的活性。结果至少重复3次。
1.2.5 实时荧光定量PCR(qRT-PCR)
使用TRIzol 试剂从细胞系中提取总RNA,然后使用PrimeScript RT Master Mix Perfect Real Time Kit将其反转录为第一链cDNA。qRT-PCR 使用SYBR Green 技术在LightCycler480Ⅱ(罗氏公司,美国)中进行。反应条件为:95 ℃10 min预变性,95 ℃15 s,60 ℃30 s,72 ℃30 s,共40 个循环,最后72 ℃延伸5 min。所有样品均以β-actin 作为标准品进行3 次重复分析,并使用2-ΔΔCt方法评估相对表达水平。具体引物列于表1。

表1 小干扰序列和qRT-PCR引物Table 1 Sequences of siRNAs and qRT-PCR primers
1.2.6 蛋白免疫印迹实验
使用试剂盒从细胞系中提取总蛋白,然后加入5×上样缓冲液并100 ℃煮沸10 min。配制10%SDSPAG凝胶,80 V预电泳后100 V恒压分离蛋白,分离蛋白后恒流250 mA,120 min 转至PVDF 膜,室温下5%脱脂奶粉溶液封闭2 h 后4 ℃孵育一抗过夜,使用TBST 洗膜后室温孵育二抗1 h,再次洗膜后加曝光液曝光显影。使用GAPDH 作为内参,实验独立重复3次。
1.3 统计学方法
采用GraphPad Prism 7及SPSS 22.0软件对数据进行统计分析。实验结果以均数±标准差()表示,两组间比较采用t检验。P<0.05为差异有统计学意义。
2 结果
2.1 人Six1启动子活性检测
将合成的pSix1-451 片段质粒进行核酸序列测定,结果提示此片段核苷酸序列与人Six1基因启动子区序列一致,说明人Six1基因启动子区已经成功插入pGL3-basic载体中。如图1所示,与pGL3-basic组相比,转染pSix1-451片段质粒的HEK-293和BEAS-2B 细胞相对荧光素酶活性明显增加,差异有统计学意义(P<0.001)。结果表明,人Six1 启动子荧光素酶报告重组质粒转染至HEK-293 和BEAS-2B细胞后具有启动子活性。

图1 荧光素酶报告基因分析HEK-293和BEAS-2B细胞中人Six1启动子活性Figure 1 Human Six1 promoter activity in HEK-293 and BEAS-2B cells analyzed by luciferase reporter system
2.2 人Six1启动子区转录因子结合位点预测
根据上述人pSix1-451 片段质粒的荧光素酶活性检测结果分析,提示-351~+100 nt 区域内可能存在明显影响人Six1 启动子活性的转录因子结合位点,应用JASPAR对该区域进行分析,获得可能的转录因子结合位点。如图2 所示,预测结果提示该序列范围内可能存在E2F4等转录因子结合位点。

图2 运用JASPAR网站预测转录因子结合位点Figure 2 Predication of binding sites of transcriptional factors by JASPAR
2.3 转录因子E2F4对Six1启动子水平的调控
为了确定E2F4能否直接调节人Six1的转录,我们在HEK-293 和BEAS-2B 细胞中分别敲低和过表达转录因子E2F4。如图3所示,与siNC处理的对照组相比,用siE2F4处理的HEK-293和BEAS-2B细胞中,pSix1-451 片段质粒荧光素酶的活性明显降低(P<0.001,P<0.01);而与用空载体pENTER 处理的对照组相比,用pE2F4 处理的HEK-293 和BEAS-2B细胞中,pSix1-451片段质粒荧光素酶的活性明显升高(P<0.05,P<0.05)。

图3 转录因子E2F4在启动子水平调控HEK-293和BEAS-2B细胞人Six1基因Figure 3 The transcription factor E2F4 regulates the human Six1 gene at the promoter level in HEK-293 and BEAS-2B cells
2.4 转录因子E2F4对Six1 mRNA水平的调控
为进一步探究E2F4对人Six1的调控作用,通过qRT-PCR 测量了HEK-293 和BEAS-2B 细胞中E2F4和Six1 的mRNA 水平,以确定E2F4 是否影响Six1 mRNA 的表达。与siNC 处理的对照组相比,siE2F4处理的HEK-293和BEAS-2B细胞中,内源性Six1的mRNA 水平明显降低(P<0.01,P<0.001,图4)。同时,与pENTER 处理的对照组相比,pE2F4 处理的HEK-293 和BEAS-2B 细胞中,内源性Six1 的mRNA水平明显升高(P<0.05,P<0.001,图4)。
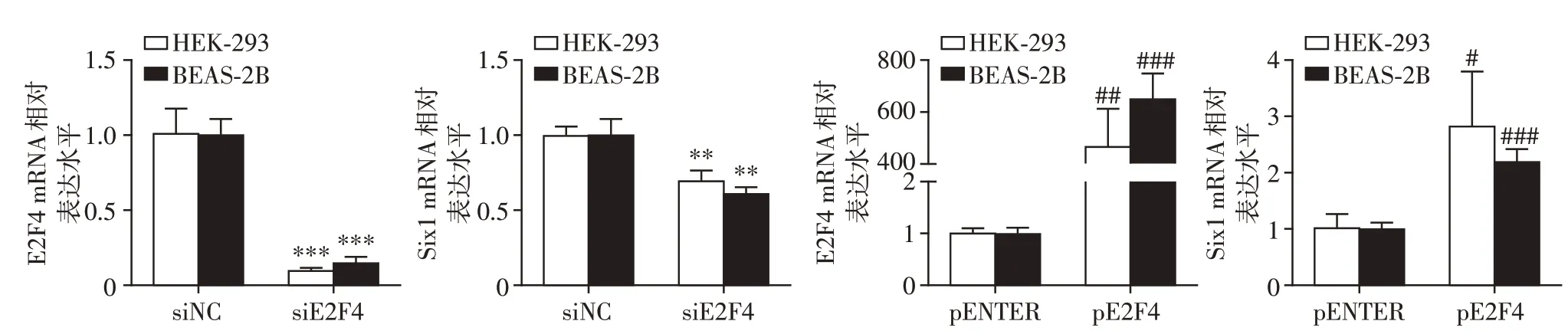
图4 转录因子E2F4在mRNA水平调控HEK-293和BEAS-2B细胞人Six1基因表达Figure 4 The transcription factor E2F4 regulates the human Six1 gene at the mRNA level in HEK-293 and BEAS-2B cells
2.5 转录因子E2F4对Six1蛋白水平的调控
通过蛋白免疫印迹实验分别检测HEK-293 和BEAS-2B细胞中敲低和过表达E2F4后E2F4和Six1的蛋白水平,以确定E2F4是否影响Six1蛋白水平的表达。如图5 所示,与siNC 处理的对照组相比,siE2F4处理的细胞中内源性Six1的蛋白水平明显降低(P<0.01)。同时,与pENTER 处理的对照组相比,pE2F4 处理的细胞中内源性Six1 的蛋白表达水平明显升高(P<0.05)。

图5 转录因子E2F4在蛋白水平调控HEK-293和BEAS-2B细胞人Six1基因表达Figure 5 The transcription factor E2F4 regulates the human Six1 gene at the protein level in HEK-293 and BEAS-2B cells
3 讨论
真核生物在其转录水平的调控是真核生物调控基因表达非常重要的机制。转录水平的调控主要通过调控转录起始部位一些特殊的调控序列,如启动子和作用于启动子区的一些转录因子。启动子作为转录的起始,指导着RNA聚合酶Ⅱ对启动的调控,对基因的正确转录起着关键作用,是蛋白质编码基因以及真核生物中许多非编码基因表达的重要组成部分,但也是真核生物转录调控中最容易被忽视的一部分[11]。对人Six1 基因的研究多集中在其蛋白水平的研究,但对其启动子的转录调控却很少有研究。本研究通过构建人Six1 启动子荧光素酶基因报告重组质粒pSix1-451,并通过瞬时转染和荧光素酶活性检测,证实其具有启动子活性。然后通过JASPAR 软件预测其可能含有E2F4 在内的多个转录因子结合位点,并在启动子、mRNA和蛋白多水平验证了转录因子E2F4 对人Six1 基因转录的正向调控。
Six家族是一组进化保守的转录因子,存在于从扁虫到人类的多种生物中,在视网膜、耳、鼻、大脑、肾脏、肌肉和性腺中的细胞群体表达,并在其中发挥关键的作用。Six1 基因是Six 基因家族中重要的一员,参与了许多组织和器官的发育,如肌肉、肾脏、听觉系统、感觉器官和颅面结构[12],并在多种肿瘤组织中高表达,如乳腺癌、肺癌、伯基特淋巴瘤、肝细胞癌、卵巢癌和横纹肌肉瘤等[13]。有研究表明,Six1 的下调直接影响气道重塑重要介质基质金属蛋白酶(matrix metalloproteinase,MMP)-2和MMP-9 的表达和分泌。此外,沉默Six1 可以显著抑制肺中核因子κB(NF-κB)途径的激活。在慢性哮喘小鼠中炎症细胞(包括嗜酸性粒细胞、嗜中性粒细胞、巨噬细胞、肥大细胞和T淋巴细胞)比在PBS处理的对照小鼠中的表达量更高,而敲低Six1显著减少了炎症细胞的积累,从而有效抑制了哮喘小鼠的气道炎症和气道重塑[14]。Six1 是由非造血细胞(如间充质系细胞)通过TGF-β、白介素(interleukin,IL)-1β、肿瘤坏死因子(tumor necrosis factor,TNF)-α、IL-6和脂多糖(lipopolysaccharide,LPS)刺激产生的。这些间充质细胞通过释放Six1作用于免疫细胞,促使其分泌促炎性细胞因子和趋化因子[15]。Six1影响靶细胞信号事件的机制一直是研究的重点,研究确定了至少5 个潜在的Six1 信号途径,包括抑制磷酸化的Smad 1、3、8 信号通路、AKT 途径、AMP 激活的蛋白激酶途径、TLR4/CD14 途径以及对Na+/K+-ATP 酶膜电位方面的影响[16-17]。因此,调控Six1自身的表达也可以通过调控有关蛋白以及各种信号通路直接或间接起作用,并为后续进一步的研究提供了方向。
转录因子是一类可以和基因上游启动子区序列结合从而调控相关基因表达的蛋白质。E2F转录家族在高级真核生物中编码一系列转录因子,并且在哺乳动物的细胞周期和DNA 合成中起着重要调控作用。目前根据其功能特征分为两类:即转录激活因子,包括E2F1、E2F2和E2F3a,以及转录阻遏因子包 括E2F3b、E2F4、E2F5、E2F6、E2F7 和E2F8[18]。E2F4作为E2F转录家族的一员,已被证实和子宫颈癌、Burkitt淋巴瘤等多种疾病相关[19],并且在细胞增殖、周期、凋亡和侵袭中发挥重要作用[20]。研究表明E2F4 的沉默减少了人类肠道上皮细胞和结直肠癌CRC细胞的增殖[21],但其在哮喘中的作用却很少有研究。
本研究首先成功构建了人Six1 启动子荧光素酶报告重组质粒,测定其具有启动子活性。其次通过软件预测其启动子区可能含有的关键转录因子结合序列。最后,在启动子、mRNA和蛋白水平验证了转录因子E2F4对人Six1的正向转录调控。
