丙酮酸乙酯通过Nrf2途径抑制谷氨酸对大鼠小脑颗粒神经元的毒性作用*
2021-01-06欧阳颖曾爱红张羚枚周瑞瑜唐淑敏阮扬皓李伍凤
欧阳颖,曾爱红,张羚枚,周瑞瑜,唐淑敏,阮扬皓,李伍凤
(中山大学孙逸仙纪念医院儿科,广东广州 510120)
新生儿缺氧缺血性脑病(hypoxic-ischemic en⁃cephalopathy,HIE)是新生儿窒息后的严重并发症,是引起智能落后、癫痫、共济失调和脑性瘫痪[1]的主要原因之一。据报道,近10年来HIE的发病率大约在(2~3)/1 000 例活产新生儿[2-3]。到目前为止,国内外对新生儿缺氧缺血性脑病的治疗,主要包括支持治疗和对症治疗,但均对已有的脑损伤无作用。因此,寻找更为有效的治疗方法成为当前急需解决的问题。
HIE 是围产期新生儿缺氧、缺血引起的脑部病变,在大多数情况下,缺氧和缺血同时发生,但缺血对中枢神经系统损伤更大。缺氧缺血性脑病发病机理极为复杂,现阶段普遍认为HIE 是代谢障碍、酸中毒、兴奋性氨基酸的毒性作用、细胞内钙超载、氧化应激反应、炎症反应等一系列病理作用的结果[4-6]。因此,明确HIE 的发病机制、寻找高效且副作用少的治疗方法具有重要的意义。
丙酮酸乙酯(ethyl pyruvate,EP)是丙酮酸的酯化物,是一种化学性能稳定、亲脂性的小分子化合物,能迅速穿过细胞膜和线粒体[7]。EP 的主要药理作用包括提供能量代谢物,保护细胞免受氧化产物和自由基的侵害,抑制炎症反应[8-11]。有报道称EP通过核因子E2 相关因子2(nuclear factor E2-related factor 2,Nrf2)/抗氧化反应元件(antioxidant response element,ARE)/血红素加氧酶1(heme oxygenase-1,HO-1)通路激活内源性抗氧化系统[10]。此外,EP 还通过与P65[11]相互作用抑制NF-κB的活性,降低早期肿瘤坏死因子α(tumor necrosis factor-α,TNF-α)和白细胞介素6(interleukin-6,IL-6)等炎症因子的表达,下调[12]晚期HMGB1的合成和释放。因此,EP 有望通过逆转HIE 的病理过程,发挥更大的神经保护作用。研究EP 作用下体外培养新生大鼠颗粒神经元Nrf2/ARE/HO-1 信号通路的激活状态并探讨其机制,将为HIE未来临床治疗提供参考资料。
材料和方法
1 主要试剂
基础细胞培养液和新生小牛血清购自Gibco BRL;胰酶、脱氧核糖核酸酶、胰酶抑制剂、阿糖胞苷、核糖核酸酶、谷氨酸(glutamate,Glu)、多聚赖氨酸、二乙酸荧光素(fluorescein diacetate,FDA)、碘化丙 啶(propidium iodide,PI)、Hoechst 33258、EP 和MK-801 均购自Sigma;硫酸庆大霉素购自福建三明制药厂;Triton X-100和氯仿购自Merck;饱和重蒸酚溶液购自中山大学医学院药理实验室;细胞核蛋白与细胞质蛋白提取试剂盒购自宁波唯奥公司;H2-DCF-DA 购自Invitrogen;抗Nrf2 和HO-1 抗体购自Stressgen;MTT 购自Sigma-Aldrich;二氢乙啶(dihy⁃droethidium,DHE)和4%多聚甲醛购自Sigma。
2 小脑颗粒神经元培养
按文献方法[13],取新生7 d 的Sprague-Dawley(SD)大鼠,雌雄不拘,在无菌条件下断头、开颅,取小脑,放在解剖液[10×Krebs 缓冲液(NaCl 72.5 g,KCl 4.0 g,NaH2PO41.4 g,右旋糖26 g,苯酚红100 mg,EP 59.6 g/L,pH 7.4)15 mL,白蛋白0.45 g,3.82%MgSO41.2 mL,pH 7.4]中,显微镜下去除血管和脑膜。然后用眼科剪剪成1 mm×1 mm×1 mm 大小的组织块,胰酶(0.25 g/L)消化(37℃,15 min),立即加反应终止液I(含胰酶抑制剂0.52 g/L、DNase-I 0.08 mg/L和Mg2+3 mmol/L 的解剖液)混匀、离心、弃上层,沉淀加反应终止液II(含Mg2+3 mmol/L 和Ca2+100 μmol/L 的解剖液)2 mL 反复吹打,离心(319×g,30 s),去除未消化的组织,细胞悬液再离心(958×g,5 min),弃上层,细胞沉淀以完全培养液(BME,25 mmol/L KCl,谷氨酰胺20 mmol/L,10%胎牛血清,0.1 g/L 硫酸庆大霉素)分散,按(1.5~1.8)×109/L 种植在预先铺有多聚赖氨酸(5 mg/L)的培养板或培养皿中,细胞数为(2~3)×106/cm2。然后置于CO2培养箱中(37℃,5% CO2)培养。24 h 后,加阿糖胞苷(终浓度为10 μmol/L)。7 d 后,加葡萄糖(终浓度为5 mmol/L)。以后每隔4 d 按前法加一次葡萄糖。7 d后,神经元分化成熟。
3 Glu兴奋性模型建立与实验分组
将体外培养7 d 的小脑颗粒神经元随机分为:(1)空白对照(control)组;(2)Glu 组[参考文献[13],弃去培养液,加入Locke 溶液(含154 mmol/L NaCl,5.6 mmol/L KCl,3.6 mmol/L NaHCO3,2.3 mmol/L CaCl2,5.6 mmol/L glucose,10 mmol/L HEPES,pH 7.4)和1 mol/L 甘氨酸,以充分激活NMDA 敏感的Glu 识别位点,Glu 浓度为100 μmol/L,30 min 后换成普通的完全培养液];(3)Glu+EP 组[在(2)组基础上加入不同浓度(0.1、0.5、1、2.5 和5 mmol/L)的EP。每组有8个复孔。
4 主要方法
4.1 FDA/PI 荧光染色 用FDA 和PI 分别对活细胞和死细胞进行染色。将待处理的细胞,弃培养液后,以含糖的PBS 洗1 遍,加入FDA(10 mg/L)和PI(4 mg/L),于暗室37℃孵育5 min,弃染液后以含糖的PBS 洗2 遍,荧光显微镜下拍照,每组随机选3 个视野拍照。与正常对照组比较,计算存活率。细胞存活率(%)=各处理组的活细胞数/正常对照组的活细胞数×100%。
4.2 MTT 实验 取培养7 d 的小脑颗粒细胞,在给药相应时间后,加入新鲜配制的MTT(溶于PBS,5 g/L)20 μL,将细胞置于含5%CO2、37℃的培养箱中孵育4~6 h后去除培养液,加入二甲基亚砜溶解结晶体,待结晶体完全溶解后,在酶标仪上(测定波长570 nm,参考波长630 nm)测吸光度(A),细胞存活率(%)=A处理组/A对照组×100%。
4.3 细胞内游离钙浓度(intracellular free calcium concentration,[Ca2+]i)的测定 用培养第7 d 的细胞,经5 μmol/L的Fluo-3/AM负载60 min,用Hanks液洗涤3次,放在倒置荧光显微镜下,由自动灌洗仪持续灌流缓冲液。应用激光扫描共聚焦显微镜观察细胞内Fluo-3 的荧光强度(激发波长488 nm,发射波长526 nm),每组选取20个活细胞,每个细胞间隔2 s扫描1 次,共扫描16 次,记录16 个荧光强度数值,取平均值为该细胞的荧光强度。根据公式计算:[Ca2+]i=Kd(F−Fmin)/(Fmax−F),Kd为Fluo3 与Ca2+反应的解离常数,在室温下为400 nmol/L;F 为实验记录到的荧光强度值;Fmax为最大荧光值,加入5 mmol/L CaCl2的EGTA 溶液所测得的值;Fmin为最小荧光值,由在无Ca2+缓冲液中加入10 mmol/L EGTA测得。
4.4 H2-DCF-DA 检测 ROS 在Glu 模型作用后90 min,弃去培养液,使用PBS 清洗3 次后,加入终浓度为30 μmol/L 的H2-DCF-DA,在37℃培养箱中孵育20 min,用PBS清洗1次,迅速置于荧光酶标仪检测荧光强度,激发及吸收波长采用475 nm/525 nm。
4.5 DHE 检测超氧阴离子 在Glu 模型作用后90 min,弃去培养液,使用PBS 清洗3 次后,加入终浓度为10 μmol/L 的DHE,在37℃培养箱中孵育30 min,用PBS清洗1次,迅速置于多功能荧光酶标仪检测荧光强度,激发及吸收波长采用485 nm/535 nm。
4.6 Western blot 根据试剂盒说明,提取细胞核蛋白与细胞质蛋白,于−80℃保存,后与DTT 混合、煮沸、离心,使蛋白质变性,取上清20 μL 于200 V 进行SDS-PAGE 45 min;取出硝酸纤维膜,浸入TBS+TBST+5%脱脂奶粉溶液,室温封闭2 h,在10 mL I 抗稀释液中把NC 膜和I抗共同轻摇孵育,4℃过夜。第2天,用1×TBST 洗NC 膜3次(每次15 mL,5 min),把NC 膜与溶解在10 mL 封闭液中的与HRP 结合的Ⅱ抗(适当比例稀释)和HRP 结合的抗生物素抗体(1∶1 000)室温下共同孵育1 h;用1×TBST 洗NC 膜3 次(每次15 mL,5 min),与10 mL 1× LumiGLO(含0.5 mL 20×底物B和9.0 mL水)室温下轻摇1 min。排去过量的液体,在暗室中使其与X 光片在增感屏中紧密接触,使X 光片感光数秒到数分钟。显影,定影,冲洗,晾干,拍照并进行计算机图像分析。
4.7 免疫组化 细胞用PBS 溶液洗2 次,后用4%多聚甲醛固定在室温下30 min。然后再用PBS 洗2次,通过0.5% Triton X-100 在室温下孵化15 min。10%的细胞封锁在山羊血清,室温1 h。4℃孵育过夜。PBS 洗涤3 次,室温下用稀释的荧光抗体孵育1.5 h。室温下用Hoechst 33258 染色细胞核15 min,PBS洗涤3次后,激光共聚焦显微镜成像。
4.8 ARE诱导转录活性检测 用Promega公司双萤光素酶报告基因试剂盒检测荧光强度。所有的程序都是按照制造商的指示进行的。pGL6-ARE-Luc 质粒是基于pGL6 载体构建,ARE 序列为3 个串联的CACCGTGACTCAGCAATT,并在我们的实验中进行。诱导转录活性增加倍数=(药物诱导的萤火虫萤光素酶活性/药物诱导的海肾萤光素酶活性)/(对照组诱导的萤火虫萤光素酶活性/对照组诱导的海肾萤光素酶活性)。
5 统计学处理
用SPSS 16.0 统计软件进行分析。以上实验均重复3次,数据均采用均数±标准差(mean±SD)表示,多组间比较采用单因素方差分析,组间两两比较采用LSD法。以P<0.05为差异有统计学意义。
3.3.1 加强电力需求侧管理。在现有用户负荷管理范围内,选取部分可中断负荷,建设毫秒级精准负荷控制系统,优化升级现有负荷管理系统。应用大数据平台,试点对用户用电行为开展实时分析,进一步挖掘其在用电预测、错峰调度等方面潜力,促进供需互动。
结 果
1 EP对小脑颗粒神经元的保护作用
如图1 所示,EP 对原代培养神经元Glu 兴奋性毒性的抑制作用呈剂量依赖性。100 μmol/L Glu 对原代培养的小脑颗粒神经元有明显的损伤作用,表现为细胞数量减少,细胞体积异常,轴突/树突断裂和消失。FDA/PI双染色下可见正常组绝大部分细胞染成绿色,夹杂着少许红染的细胞,Glu 组则绿染细胞减少,而红色的死细胞明显增多。1 mmol/L EP 预处理30 min 后,Glu 诱导损伤下存活神经元数量显著增加(P<0.05),EP以浓度依赖性的方式显著提高了神经元的活力。
2 细胞内ROS检测结果
我们通过H2-DCF-DA 检测小脑颗粒神经元的ROS 含量,用DHE 检测超氧阴离子含量。H2-DCFDA 染色后,含ROS 的细胞发绿色荧光,ROS 越多,荧光强度越大。与正常对照组少量的绿色荧光细胞相比,Glu 组可见到绿色荧光细胞明显增多,荧光强度大;而Glu+EP 组则绿色荧光细胞明显减少,荧光强度明显减弱,见图2。DHE 染色后,含超氧阴离子的细胞与DHE 作用而发出红色荧光,超氧阴离子越多,荧光强度越大。与正常对照组少量红色荧光细胞相比,Glu 组可见红色荧光细胞明显增多,荧光强度增大;而Glu+EP 组则红色荧光细胞明显减少,荧光强度较弱,见图2。
3 小脑颗粒神经元[Ca2+]i检测结果
我们通过Fluo-3 作为指标检测细胞内Ca2+。在小脑颗粒神经元中,加入Glu,动态观察[Ca2+]i水平,20 s 即可观察到荧光强度迅速升高,达到高峰,随时间延长逐渐降低,但6 min 时仍显著高于正常;加入EP(2.5 和5.0 mmol/L)预处理30 min 可以部分阻滞[Ca2+]i升高(P<0.05),见图3。
4 EP通过激活Nrf2/ARE/HO-1通路发挥神经保护作用
通过检测EP 处理神经元中Nrf-2 的表达,观察到EP 组中细胞质Nrf2 表达下降,细胞核Nrf2 表达明显上调,见图4A。之后,在Nrf2 免疫组化染色后加入Hoechst 染色,在激光共聚焦显微镜下观察,正常对照组细胞浆染成红色,核染成蓝色,而EP 组中细胞质染成红色,细胞核内因Nrf2明显升高,核亦染成红色,同时在Hoechst 染色后呈蓝色,融合后核呈红色,提示Nrf2 转入核内,见图4B。在检测ARE 的转录活性实验中,EP 组ARE 的转录活性显著上调(P<0.05),见图4C。如图4D 所示,Glu 或EP 处理3 h、6 h 和12 h 均显著提高HO-1 的表达水平(P<0.05),且Glu 和EP 共处理进一步增加了HO-1 的表达。相反,HO-1 抑制剂锌原卟啉(zinc protoporphyrin,ZnPP)部分抑制了EP的神经保护作用,见图4E。
讨 论
1 Glu对体外培养小脑颗粒神经元的毒性作用
本实验中,小脑颗粒神经元在Glu作用后24 h存活率为(28.4±2.1)%,与文献结果相符[14]。DNA 电泳未出现梯形条带,呈涂布状,提示主要的死亡方式为坏死。另一研究[15]的结果则不同,在没有甘氨酸存在及无镁离子的Locke 液的情况下,Glu 的毒性浓度选择为300 μmol/L,但细胞存活率仍高于本实验模型。因此,同样为Glu模型,浓度不同,处理方式不同,细胞死亡的方式不同,Glu 引起细胞死亡所激活的通路可能不同。
2 EP对小脑颗粒神经元的保护作用
EP 具有组织能量守恒、抗炎、抗氧化等药理作用[16],可减轻体内的全身炎症反应和多脏器功能障碍[17]。有研究称EP 可以治疗重症急性胰腺炎(se⁃vere acute pancreatitis,SAP),减轻SAP 相关的远端器官损伤,如肝、肺和肾[18-19]。此外,EP 能减轻外伤性脑损伤和出血性脑损伤[20]。
在本研究中,EP在浓度0.1~5 mmol/L时剂量依赖性地抑制Glu 的兴奋性毒性作用,充分说明EP 具有神经保护作用,可能为一种在中枢神经系统疾病中有应用前景的药物。
3 细胞内ROS检测结果
4 EP抑制细胞内钙升高
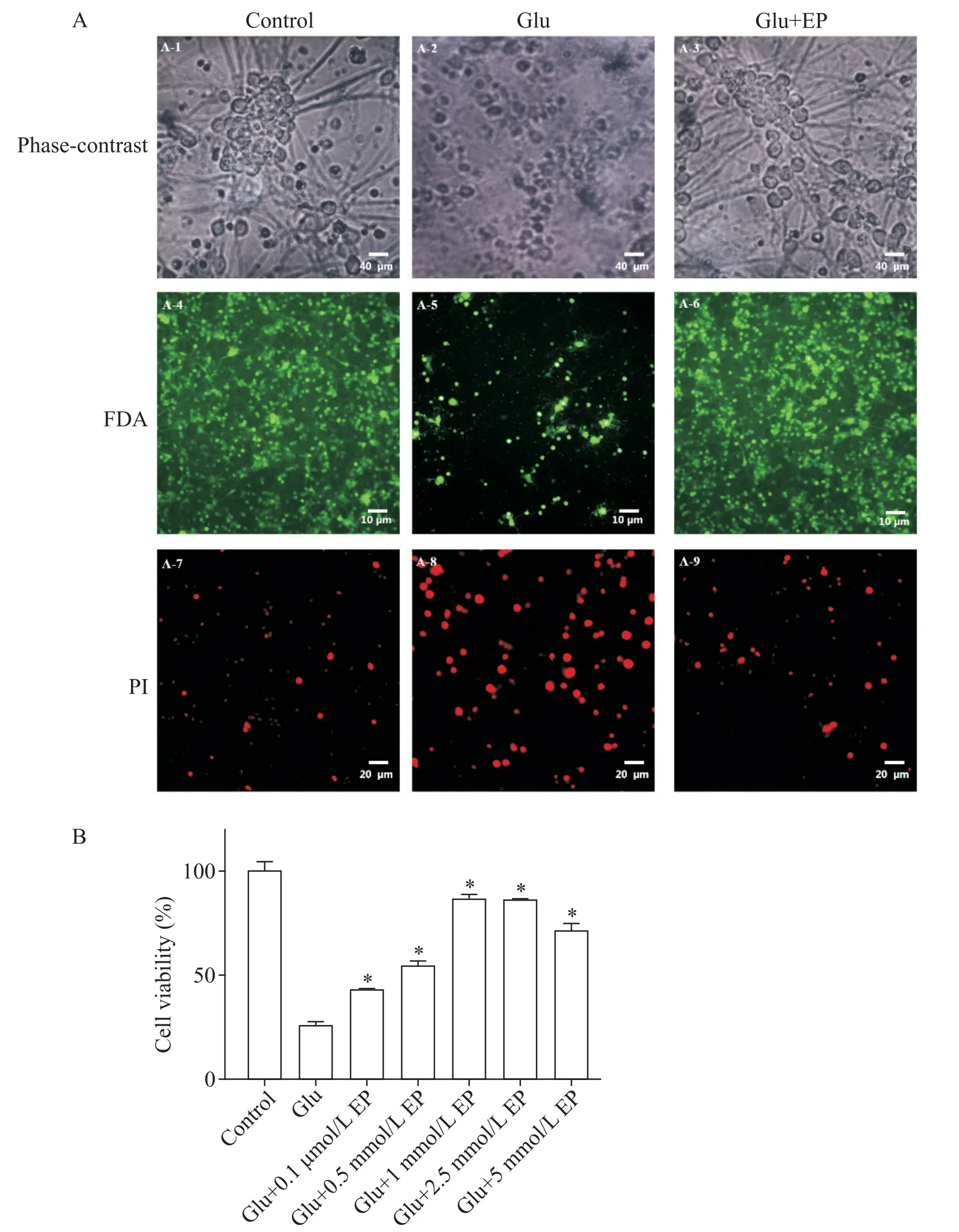
Figure 1.Protective effect of ethyl pyruvate(EP)on rat cerebellar granule neurons.A-1:normal cerebellar granule neurons,scale bar=40 μm;A-2:cerebellar granule neurons treated with 100 μmol/L glutamate(Glu)for 24 h,scale bar=40 μm;A-3:cerebellar granule neurons treated with Glu(100 μmol/L)+EP(2.5 mmol/L)for 24 h,scale bar=40 μm;A-4:normal cer⁃ebellar granule neurons,FDA staining,scale bar=10 μm;A-5:cerebellar granule neurons treated with Glu(100 μmol/L)for 24 h,FDA staining,scale bar=10 μm;A-6:cerebellar granule neurons treated with Glu(100 μmol/L)+EP(2.5 mmol/L)for 24 h,FDA staining,scale bar=10 μm;A-7:normal cerebellar granule neurons,PI staining,scale bar=20 μm;A-8:cerebellar granule neurons treated with Glu(100 μmol/L)for 24 h,PI staining,scale bar=20 μm;A-9:cerebellar granule neurons treated with Glu(100 μmol/L)+EP(2.5 mmol/L)for 24 h,PI staining,scale bar=20 μm.B:the effect of EP(0.1~5 mmol/L)on the viability of Glu(100 μmol/L)-induced cerebellar granule neurons.Mean±SD. n=3.*P<0.05 vs Glu group.图1 丙酮酸乙酯对大鼠小脑颗粒神经元的保护作用
正常情况下,细胞内的钙浓度为100 nmol/L,比细胞外的钙低10 000倍。细胞间隙高浓度的Glu,可引起N-甲基-D 天冬氨酸受体过度兴奋,Ca2+大量内流,[Ca2+]i短暂升高;经过一段时间之后,出现第2次钙离子升高,称之为延迟性钙失调,为不可逆损伤,导致细胞死亡。[Ca2+]i过高可聚集在线粒体内,损伤氧化磷酸化通道,造成ATP 不足;另一方面,Ca2+与钙调素形成复合物,激活多种蛋白酶,引起广泛的信号传导分子磷酸化,而产生各种细胞效应;同时促进氧自由基产生,故细胞内钙超载被认为是细胞死亡的最后通路[22]。
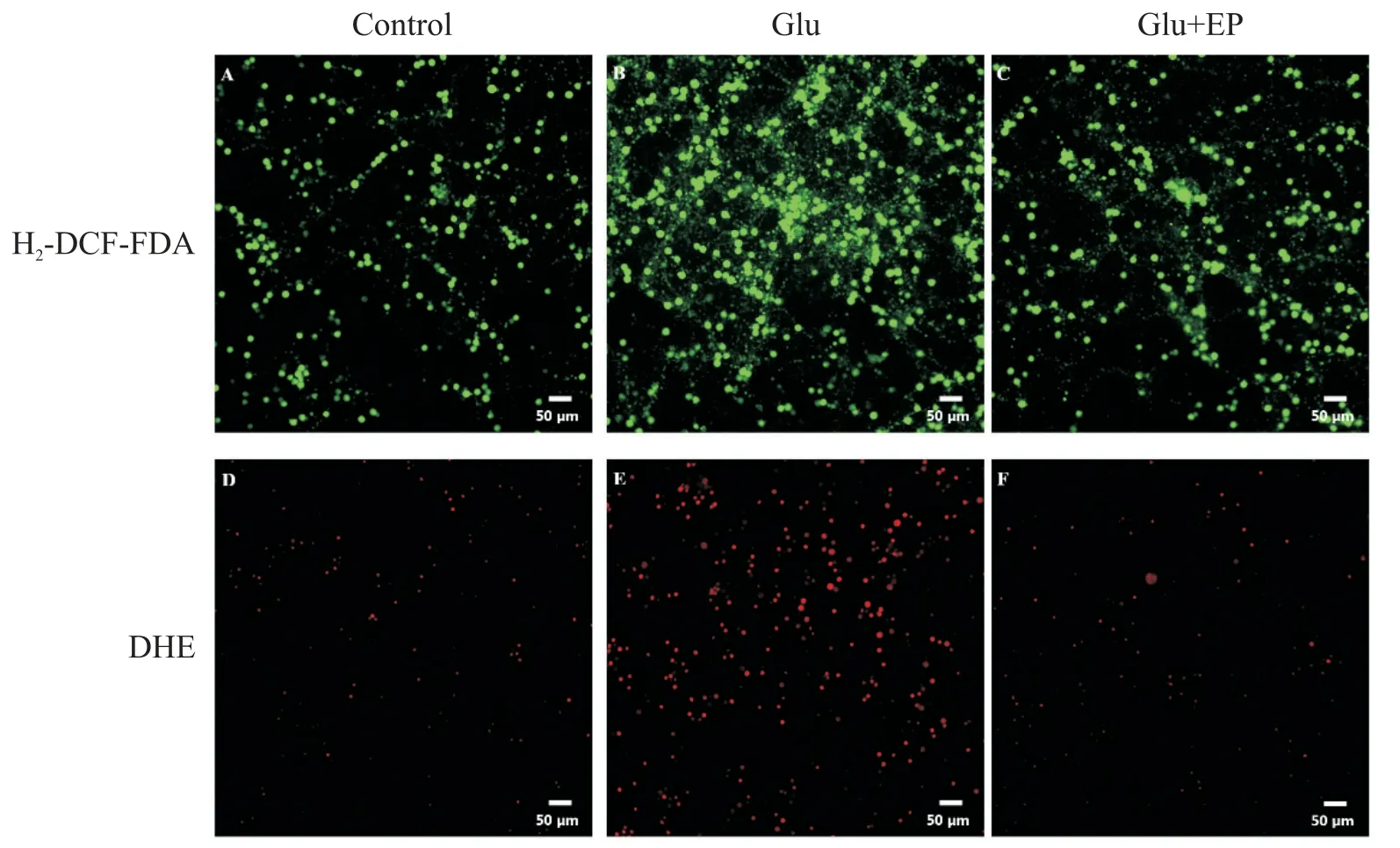
Figure 2.Detection of ROS in rat cerebellar granule neurons.A:control group,H2-DCF-DA staining;B:glutamate(Glu;100 μmol/L)group,H2-DCF-DA staining;C:Glu(100 μmol/L)+ethyl pyruvate(EP;2.5 mmol/L)group,H2-DCF-DA stain⁃ing;D:control group,DHE staining;E:Glu(100 μmol/L)group,DHE staining;F:Glu(100 μmol/L)+EP(2.5 mmol/L)group,DHE staining.Scale bar=50 μm.图2 大鼠小脑颗粒神经元内ROS检测结果
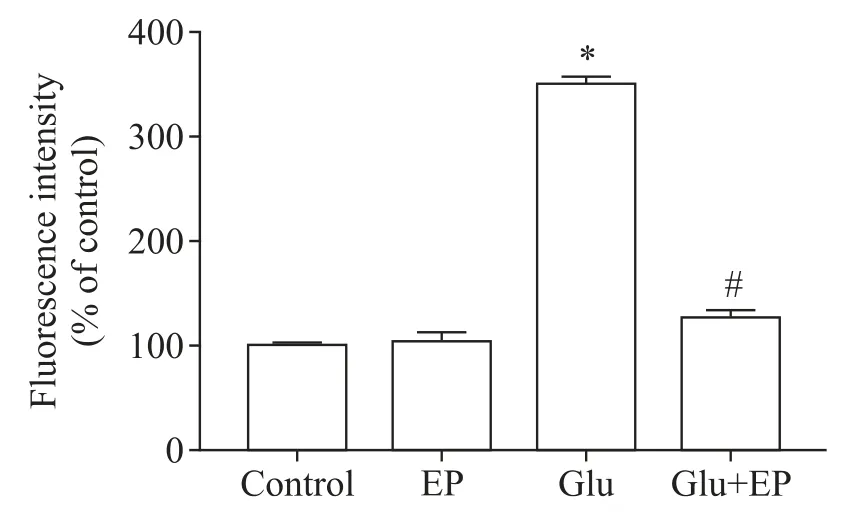
Figure 3.Detection of[Ca2+]i in rat cerebellar granule neurons.Mean±SD. n=3.*P<0.05 vs control group;#P<0.05 vs Glu group.图3 大鼠小脑颗粒神经元[Ca2+]i检测结果
本实验中,Glu 组细胞内钙在20 s 左右迅速升高,持续6 min 以上。而EP+Glu 组细胞内钙较正常组升高,但显著低于Glu组,提示EP通过抑制细胞内钙离子浓度升高,阻断了引起神经元损伤的上游通路。因此,抑制[Ca2+]i的升高在EP 的神经保护作用中起到很重要的作用。
5 EP 通过激活Nrf2/ARE 通路而发挥神经保护作用
Nrf2-ARE信号通路被认为是一种新型的抗氧化信号通路,以Nrf2为核心,激活一些保护性的酶的产生,如HO-1、超氧化物歧化酶及还原型谷胱甘肽;HO-1 及还原型谷胱甘肽可抑制细胞凋亡,抗氧化,缓解细胞内钙离子超载[23]。在颅脑外伤、颅内出血和脑缺血梗死动物模型中均有Nrf2-ARE 信号通路激活。有研究显示,移植Nrf2 过度表达的星形胶质细胞至纹状体可减轻丙二酸诱导的细胞损害[24];应用小干扰RNA 降低Nrf2 的表达,可引起NO 诱导的神经细胞瘤细胞的凋亡增多[25]。
本实验结果显示,在EP 处理组,Nrf2 转运入核中,12 h 核内Nrf2 仍升高。免疫组化染色显示核内Nrf2 明显增多,ARE 介导的荧光素酶转录活性升高,提示EP 激活Nrf2/ARE 通路,使ARE 转录活性增高。同时,Glu 引起HO-1 升高,证明Glu 的氧化性损伤也可引起HO-1 升高,但12 h 即下降,说明Glu 激发的HO-1 升高所起到的保护作用有限,而EP 可引起正常小脑颗粒细胞内HO-1升高,与Glu一起,协同诱导HO-1升高,可发挥较强的抗氧化损伤作用。ZnPP部分阻滞EP 的保护作用,则说明HO-1 的保护作用是确切的。因此,EP 是通过激活Nrf2/ARE/HO-1 通路而起到保护作用。
综上所述,我们的研究为EP 在体外对大鼠小脑颗粒神经元Glu 兴奋性神经毒性的保护作用提供了实验依据。EP 的作用机制包括抗氧化、减轻脑水肿、抑制细胞凋亡,而在HIE 发病过程中,Glu 兴奋性毒性是脑损伤的主要环节,因此我们推断EP 可能在HIE神经保护治疗方面具有广阔的应用前景。
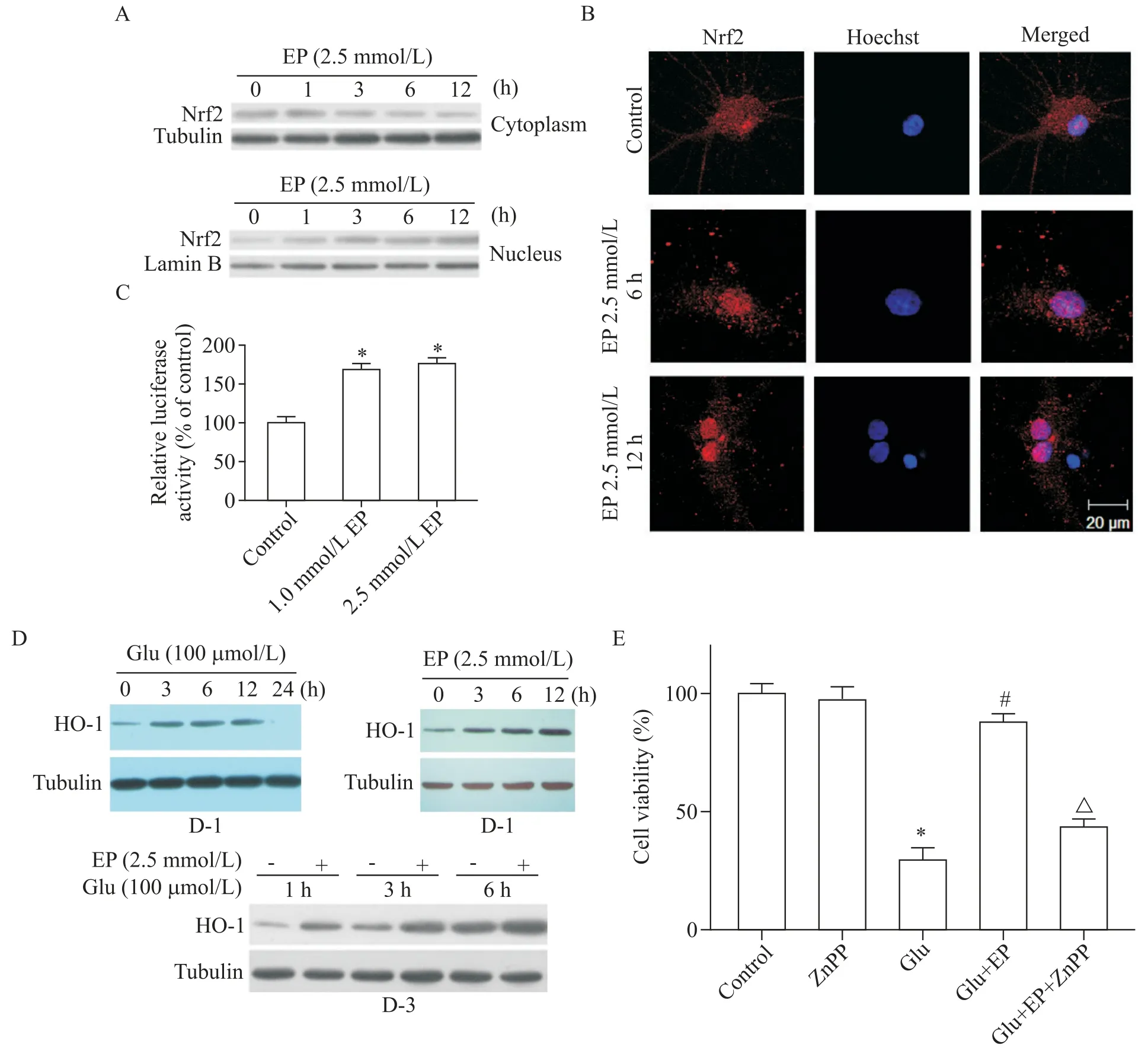
Figure 4.The expression of Nrf2/ARE/HO-1 pathway-related proteins in rat cerebellar granule neurons.A:ethyl pyruvate(EP;2.5 mmol/L)decreased cytoplasmic Nrf2,but increased nuclear Nrf2;B:cytoplasmic Nrf2 was translocated into the nucleus after treatment with 2.5 mmol/L EP for 6 and 12 h(scale bar=20 μm);C:EP elevated the relative luciferase activity in⁃duced by ARE;D-1:the expression of HO-1 after treatment with 100 μmol/L glutamate(Glu)for 0,3,6,12 and 24 h;D-2:the expression of HO-1 after treatment with 2.5 mmol/L EP for 0,3,6 and 12 h;D-3:the expression of HO-1 after treatment with 2.5 mmol/L EP+100 μmol/L Glu for 1,3 and 6 h;E:ZnPP partly inhibited the protective effect of EP against Glu excitotoxicity.Mean±SD. n=3.*P<0.05 vs control group;#P<0.05 vs Glu group;△P<0.05 vs Glu+EP group.图4 大鼠小脑颗粒神经元内Nrf2/ARE/HO-1通路相关蛋白表达的检测结果
