布鲁氏菌的胞内生存机制研究进展
2021-06-24马忠臣胡瑞瑞李芮芮努尔赛力克努素甫陈创夫
马忠臣,胡瑞瑞,李芮芮,王 震,努尔赛力克·努素甫,王 勇*,陈创夫*
(1.石河子大学动物科技学院,新疆石河子 832000;2.西部地区高发人兽共患传染性疾病防治协同创新中心,新疆石河子 832000;3.石河子大学生命科学学院,新疆石河子 832000;4.伊犁哈萨克自治州新源县农业农村局,新疆伊犁州 835800)
布鲁氏菌属(Brucella)的病原菌属于α-变形菌(Alphaproteobacteria)的α-2子群,其包含动物、植物病原体或共生菌体等多种细菌,这些细菌与真核宿主之间经历了长期的共同进化,很多生物学特性已经发生了改变[1]。目前,布鲁氏菌属的成员越来越多,几乎可以感染所有的哺乳动物,其中包括牛(Brucellaabortus),羊(B.melitensis),猪(B.suis),绵羊(B.ovis),骆驼、麋鹿、野牛(B.abortus),犬(B.canis),啮齿动物(B.neotomae和B.microti),猴子(B.papionis)和海豹、海豚、鲸鱼等海洋哺乳动物(B.pinnipedialis和B.ceti)以及两栖动物(B.inopinata)[2]。布鲁氏菌在宿主体内进化形成了一系列机制满足其胞内长期寄生与繁殖,藏匿于布氏小体(BCV)躲避早期宿主免疫杀伤是其胞内生存重要的一步,但其详细生存机制还需进一步研究。本文详细介绍了布鲁氏菌细胞内循环过程和相关分子机制,包括从eBCV到rBCV,再到aBCV的整个过程中布鲁氏菌通过伪装逃避、调节宿主分泌等满足其自身生存,这对布鲁氏菌胞内生存机制的研究具有重要意义。
1 布鲁氏菌病
布鲁氏菌(Brucella)感染会引起宿主的布鲁氏菌病(Brucellosis),在动物中表现为流产、不育和跛行,人类可以通过吸入气溶胶化的细菌、摄取、接触被污染的组织或其衍生物而感染。山羊种(B.melitensis),猪种布鲁氏菌(B.suis)和牛种布鲁氏菌(B.abortus)对人具有高致病性,此外,犬种布鲁氏菌(B.canis)和啮齿动物种布鲁氏菌(B.neotomae)也会对人造成感染[3]。人感染布鲁氏菌后会在急性期出现非特异性的类流感症状,如不及时采取抗生素治疗,将会导致慢性感染,并最终引起反复发烧、骨髓炎、关节炎、神经系统症状和心内膜炎等全身衰弱性疾病。布鲁氏菌感染在动物和人细胞水平上具有相同的病理和生理性特征,通过细胞内循环来确保其存活、增殖和寄生(包括巨噬细胞和树突状细胞)[4]。流产布鲁氏菌(Brucellaabortus)最初在感染动物的胎盘中被发现,在上皮细胞和吞噬细胞感染模型中表现出了强的增殖能力,在研究布鲁氏菌细胞内循环增殖的特性方面发挥了重要作用。
2 VirB Ⅳ型分泌系统
VirB Ⅳ型分泌系统(T4SS)的表达是布鲁氏菌毒力发挥的标志,布鲁氏菌T4SS是通过与植物病原农杆菌的VirB T4SS同源鉴定而得来,研究发现,慢性布鲁氏菌病的鼠模型以及牛、山羊和猪布鲁氏菌的细胞模型中布鲁氏菌复制必需T4SS的表达[5]。大量研究表明,VirB操纵子内多种基因的缺失或插入会抑制布鲁氏菌eBCV转化为rBCV,进而抑制布鲁氏菌在宿主细胞内的复制,也无法在小鼠中建立布鲁氏菌慢性感染模型[6],表明T4SS在rBCV生成和布鲁氏菌胞内增殖中具有重要作用。在许多胞内寄生菌中,T4SS可以分泌传递效应蛋白或核蛋白复合物,从而其发挥毒力作用,类比于那些使细菌发挥毒力的蛋白[7]。科学家推测布鲁氏菌的VirB T4SS同样可以将效应蛋白转运到宿主细胞中,跨过eBCV膜调节特定的细胞功能并介导rBCV的生成。后来,一系列分泌到宿主细胞中发挥作用的布鲁氏菌T4SS依赖性效应蛋白的发现证明了这一假设[8],为了解布鲁氏菌细胞内循环的潜在分子机制开辟了蹊径。
3 布鲁氏菌胞内循环过程
布鲁氏菌一旦入侵吞噬或非吞噬细胞,就会藏匿于膜结合隔室内,该隔室被称为布鲁氏菌小体,简称布氏小体(Brucella-containing vacuole,BCV)(图1),布氏小体是基于原始吞噬体的概念,指细菌特异的膜结合小泡。早期通过显微镜观察发现,起初的BCV通常会沿内吞途径行进,依次获取早期和后期的内体膜标记物,后被酸化,pH达到4.5[9],这类似于完整的吞噬体成熟过程。基于布氏小体在感染细胞周期(感染后0至8 h)中具有内体性质,故将其称为内体BCV(eBCV)(图1)[10]。
eBCV与内吞区室相互作用后,小泡逐渐丧失内体标记物(感染后8至12 h),并与内质网(ER)结构持续相互作用,并最终获得与ER膜相关的标记物,如钙网蛋白、钙结合蛋白和Sec61β蛋白[11],这表明该过程伴随着eBCV膜的反转和ER衍生膜的积聚。另外,这些含菌小泡还拥有了ER的结构和功能特征[11],这进一步表明eBCV已转化为ER衍生的细胞器。这些结构和功能的改变提供了促进布鲁氏菌生长和复制的胞内环境,有利于布鲁氏菌在宿主细胞内的复制。这些含菌小泡被称为复制性BCV(rBCV)(图1)[10]。
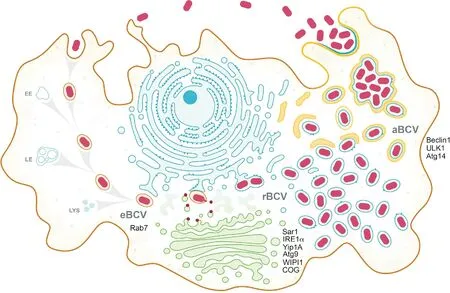
图1 布鲁氏菌在巨噬细胞内的循环模型[2]
观察rBCV的原始超微结构发现,这些小泡是单个存在的,在巨噬细胞或HeLa细胞中很少与ER相连[11],而有研究则在滋养层细胞ER腔内观察到该结构,故rBCV在宿主细胞中的确切定位尚不清晰。近年,Sedzicki J等[12]利用电子成像技术重建rBCV的三维结构,发现rBCV与ER相连,并且可能也与ER和高尔基体之间的转运囊泡相互作用,表明了rBCV与宿主分泌途径的紧密相互作用。布鲁氏菌的增殖与其rBCV阶段(感染后12 h至48 h)紧密相关,该阶段会引起宿主细胞ER迅速重组形成rBCV网[11]。在布鲁氏菌大量繁殖后,预示着布鲁氏菌胞内循环的另一个阶段开始,新月状膜结构逐渐包围吞噬rBCV,形成类似自噬体的有膜空泡,这些空泡具有后期溶酶体的特征[10]。这些重组的rBCV,称为自噬BCV(aBCV)(图1),其在感染后48至72 h形成,引发布鲁氏菌从感染细胞中释放并侵染新的细胞,aBCV的形成加速了布鲁氏菌细胞内循环的过程[10]。
布鲁氏菌在宿主细胞内由eBCV到rBCV再到aBCV三个阶段的连续循环具有功能和空间结构的多样性,它们是布鲁氏菌持续感染的必要条件。20多年来,人们对布鲁氏菌在这些阶段中潜在的分子机制做了大量的的研究,试图阐明其如何利用相应的细胞途径来完成其细胞内循环。
3.1 eBCV阶段
被吞噬细胞吞噬或侵入非吞噬细胞后,布鲁氏菌最初位于吞噬体中,并迅速获取早期内体标志物,包括控制早期内体成熟的小GTPase Rab5和早期内体抗原EEA-1[11]。由于这些标记物的获取途径与典型的吞噬体相似,因此早期BCV可能会经历一段成熟的过程。与此相一致,BCV随后获得晚期内体标志物,例如溶酶体相关膜蛋白LAMP1和LAMP2、CD63、以及控制晚期内吞体与溶酶体融合的小GTPase Rab7[9],这也表明了布鲁氏菌感染后1 h内BCV的成熟过程。重要的是,BCV也会被迅速酸化,pH达到4.5~5.0[9],这进一步说明了BCV成为吞噬溶酶体的成熟过程。 然而,通过免疫染色法检测布鲁氏菌感染早期溶酶体内容物向BCV转运来证明BCV与溶酶体的融合却未能观察到组织蛋白酶D(cathepsin D)的集聚[9]。因此,研究者提出了这样的猜想,尽管BCV已经成熟,但仍会避免与末端溶酶体融合,从而防止布鲁氏菌被免疫杀伤确保其胞内存活。
几项针对eBCV内布鲁氏菌生存能力的研究表明,由于eBCV内存在杀菌环境,大部分(原代骨髓巨噬细胞中高达90%)进入宿主细胞的布鲁氏菌被杀死[9]。Starr T等[9]使用活细胞成像技术荧光标记溶酶体检测可溶性溶酶体抗原,发现HeLa细胞中的eBCV虽与末端溶酶体融合,但程度有限,表明eBCV逐渐成熟为类吞噬溶酶体的膜泡结构。体外细胞模型中观察到eBCV成熟同样抑制了布鲁氏菌的生存增殖能力,这有悖于人们对布鲁氏菌细胞内存活的常规认识。因此有人猜测,eBCV经历完整的溶酶体成熟过程不利于布鲁氏菌的胞内存活,但也不代表其完全被杀死,其中一小部分未沿着内吞途径成熟的BCV产生rBCV来完成菌体增殖和细胞内存活。但是,相关研究表现结果与这种猜想并不太一致。首先,含有LAMP1 的eBCV与ER相互作用后获得ER标记物,暗示eBCV经历了向rBCV的转化[11]。其次,eBCV酸化至末端溶酶体的pH值是rBCV的生成和布鲁氏菌复制所必需,因为抑制溶酶体酸化会阻止eBCV向rBCV的转化和布鲁氏菌的增殖[9,13]。第三,抑制Rab7的活性可抑制eBCV与溶酶体的融合,阻碍eBCV转化为rBCV和布鲁氏菌菌增殖的过程[9],表明布鲁氏菌后期需要功能性的内吞和溶酶体融合途径完成其细胞内循环。第四,eBCV酸化对于诱导VirB T4SS表达至关重要[9],eBCV成熟后T4SS迅速表达分泌,并在感染后4 h达到高峰[14]。综上所述,eBCV阶段是布鲁氏菌细胞内循环的必要步骤,该阶段提供了诱导VirB T4SS表达和最终eBCV转化为rBCV所必需的条件,如溶酶体pH。
eBCV除了诱导布鲁氏菌T4SS的表达外,还可以促进复制性rBCV生成之前的布鲁氏菌的生长。 Deghelt T M等[15]建立的荧光显微镜方法观测到eBCV内含有LAMP1的布鲁氏菌在感染后8 h(即eBCV到rBCV的转化阶段)染色体开始复制,而G1阻滞期在菌体感染后6 h已经开始。是由eBCV的内体条件还是由转化为rBCV期间的菌小泡内变化促发了这种宿主细胞的周期性变化仍值得研究。以上这些发现表明布鲁氏菌对BCV内环境异常敏感,并且在eBCV阶段就已经开始生长,进一步强调了内体阶段在布鲁氏菌细胞内循环中的重要性。
3.2 rBCV阶段
3.2.1 ER在rBCV生成过程中的作用 rBCV的生成是布鲁氏菌细胞内循环的重要标志,其将原始的内体含菌小泡转化为可促进菌体增殖的ER来源的特殊细胞器。最初通过观察rBCV的超微结构发现,rBCV与ER融合并附有核糖体,后通过免疫荧光显微镜观察、葡萄糖-6磷酸酶活性的膜内检测进一步证实了其膜上含有ER相关蛋白。葡萄糖-6磷酸酶是一种ER腔内酶,对ER空泡毒素溶血素(ER-vacuolating toxin aerolysin)敏感[11]。BCV与晚期内吞和溶酶体区室相互作用可能发生于ER出口位点(ERES),ERES是ER子室,通过形成COPⅡ外载物囊泡(COPⅡ-coated cargo vesicles)而引发分泌转运,其功能的发挥对rBCV生成很重要[16]。在eBCV到rBCV转化过程中,依赖于T4SS与ERES COPⅡ外载物囊泡的持续相互作用(图2),并且抑制调控COPⅡ外套组装以及ERES的形成和功能的小GTPase Sar1的表达,从而抑制了rBCV的生成和布鲁氏菌增殖[16]。与这些结果相一致,另有研究发现布鲁氏菌感染后通过未知机制上调了Sar1和COPⅡ组分Sec23和Sec24D的表达量[17],表明布鲁氏菌通过上调Sar1和COPⅡ介导的分泌囊泡的运动促进rBCV生成。因此,布鲁氏菌可能通过增强分泌小泡的产生来调节ERES功能,从而促进rBCV的生成,而分泌小泡的产生可能与eBCV融合,将其转化为具有ER融合特性的小泡(图2),布鲁氏菌可能因此绕开典型的内体-高尔基体-内质网过程而直接进入内质网。
3.2.2 UPR在布鲁氏菌细胞内循环中的作用 另一种ER介导的与布鲁氏菌细胞内循环密切相关的功能是未折叠蛋白反应(unfolded-protein response,UPR)。当ER内的错误折叠蛋白积聚触发ER应激时,ER上的UPR受体激活IRE1α、PERK、ATF6信号通路,并导致ER功能重组,增强ER的折叠能力,从而抑制蛋白质合成以及引发自噬,以缓解ER应激[18]。尽管对UPR过程还存在一定的分歧,但共识是布鲁氏菌感染至少通过IRE1α途径在巨噬细胞或HeLa细胞中诱导UPR[17,19]。此外,研究表明,IRE1α是布鲁氏菌复制所必需的,牛磺酸去氧胆酸对蛋白质错误折叠的治疗作用会抑制山羊种布鲁氏菌的复制[19],这说明UPR有利于布鲁氏菌的细胞内循环。但是,这种应激反应是否以及如何促进rBCV生成和布鲁氏菌的复制尚不清楚。Taguchi Y等[170]通过展示ERES定位蛋白Yip1A激活IRE1α的过程,揭示了UPR诱导、ERES和rBCV生成之间的联系,Yip1A蛋白以依赖自噬相关蛋白Atg9和WIPI1的方式调控rBCV生成所必需的大型囊泡的形成。因此,这些发现说明布鲁氏菌可能通过IRE1α途径的激活而主动引起UPR从而促进激活级联反应,导致自噬特性的囊泡在ERES的形成,最终引起rBCV的生成(图2)。
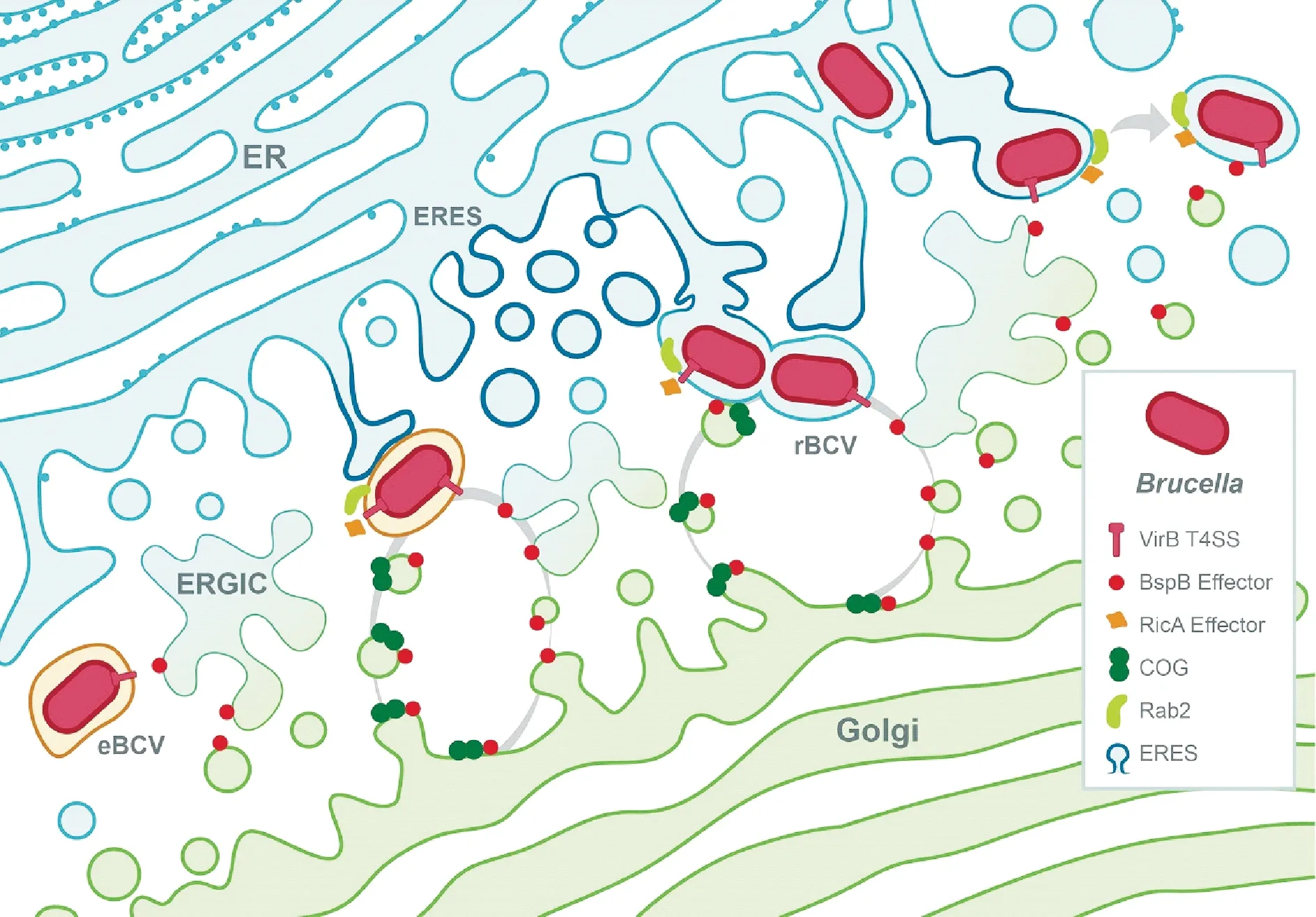
图2 rBCV的VirB T4SS依赖性生物发生模型[2]
3.2.3 自噬是否参与了rBCV的生物发生 自噬(autophagy)是一种基于膜的过程,通常将细胞内容物(胞浆内容物、受损的细胞器、微生物等)捕获到称为自噬小体的双膜囊泡中,并将其降解至溶酶体。虽然它具有先天性的免疫抗菌作用,但对某些病原体来说该过程可能是有益的。基于IRE1α和Yip1A在rBCV生成和布鲁氏菌增殖中的作用,在ERES处形成的自噬小体可能提供ER衍生的膜,从而有助于eBCV到rBCV的转化。然而,是否正常的自噬级联反应参与rBCV生物发生仍存在争议。虽然最初提出自噬过程参与了HeLa细胞中rBCV的生成,BCVs中观察到多膜结构和溶酶体复合物单丹酰尸胺(monodansylcadaverine)的聚集[20],但自噬结构在巨噬细胞eBCVs中并未见到[11]。另外,自噬标志物LC3也不在eBCV上积聚,这有悖于典型的自噬过程。此外,通过小干扰RNA(siRNA)抑制自噬因子(ULK1、beclin1、Atg5、Atg7、ATG16L和LC3B)参与的自噬体成核和延伸步骤,不能阻止rBCV的生成和布鲁氏菌的复制[9]。因此,没有实质性证据证明自噬级联反应在rBCV生成中的作用。不过,自噬体成核蛋白WIPI1和自噬蛋白Atg9在rBCV生成中的作用已被证实[17],暗示在rBCV生成过程中应用了特定的自噬相关机制,这表明ERES和Atg9介导的膜传递过程和自噬体的形成过程介导了rBCV的生成。如要阐明自噬相关过程如何真正的促进rBCV生成,还需要更深一步的研究。
3.2.4 宿主细胞分泌途径在rBCV生成中的作用 由于rBCV具有ER性质,因此布鲁氏菌细胞内循环中宿主细胞分泌途径必然起着至关重要的作用。除ER外,ER-高尔基体中间区、高尔基体和反式高尔基体网(trans-Golgi network)是否对布鲁氏菌有重要作用,还是一个值得探究的问题。研究表明,其他胞内病原体可以充分利用这些细胞器室的功能,例如嗜肺军团菌(Legionellapneumophila)通过阻逆依赖于Arf1和Rab1的分泌途径来获取ER衍生膜[21],沙眼衣原体(Chlamydiatrachomatis)重定向高尔基体脂质分泌途径以获取鞘脂(sphingolipids)[22]。通过药物抑制宿主细胞分泌途径发现,小GTP酶的显性等位基因失活或siRNA沉默抑制ER-高尔基体分泌途径,进而抑制rBCV的生成和布鲁氏菌增殖[23]。另外,使用布雷菲德菌素A(brefeldin A)或siRNA抑制Arf1分泌途径,rBCV的生成减弱至与Sar1基因缺失时相同的水平。ER-高尔基体之间转运囊泡所分泌的小GTPases Rab1a和Rab2的缺失阻碍了巨噬细胞中流产布鲁氏菌的复制[23],这均表明在eBCV转化为rBCV的过程中,这些细胞器室之间复杂的囊泡分泌可能促进布鲁氏菌的增殖。同样,与布鲁氏菌利用宿主细胞分泌途径相一致,宿主细胞感染布鲁氏菌后分泌转运途径以依赖T4SS的方式发生改变[24],这表明布鲁氏菌通过分泌效应子来主动调节宿主分泌功能以促进其细胞内循环。
3.2.5 rBCV生成过程中的布鲁氏菌效应蛋白 近年发现了15种VirB T4SS效应子(表1),其中一些可以从分子水平开始解读rBCV生成机制和布鲁氏菌增殖过程。VceC处于VirB T4SS核心位置,结合ER伴侣蛋白Grp78/BiP并诱导UPR,引发炎症反应[25],但尚不清楚它是否在rBCV生成中发挥作用。TcpB/BtpA/ Btp1,一种含有TIR(Toll/interleukin-1 (IL-1) receptor)域的效应子,可下调促炎症反应因子的表达[26-27],抑制炎症反应的发生,一旦布鲁氏菌形成rBCVs,也会诱导UPR并促进菌体增殖[19],但它可能不参与rBCV的生成过程。与之不同的是,SepA可能参与rBCV的生成过程,因为布鲁氏菌ΔsepA缺失突变体仍保留在eBCV中,无法生成rBCV[8],但其作用方式仍然未知。
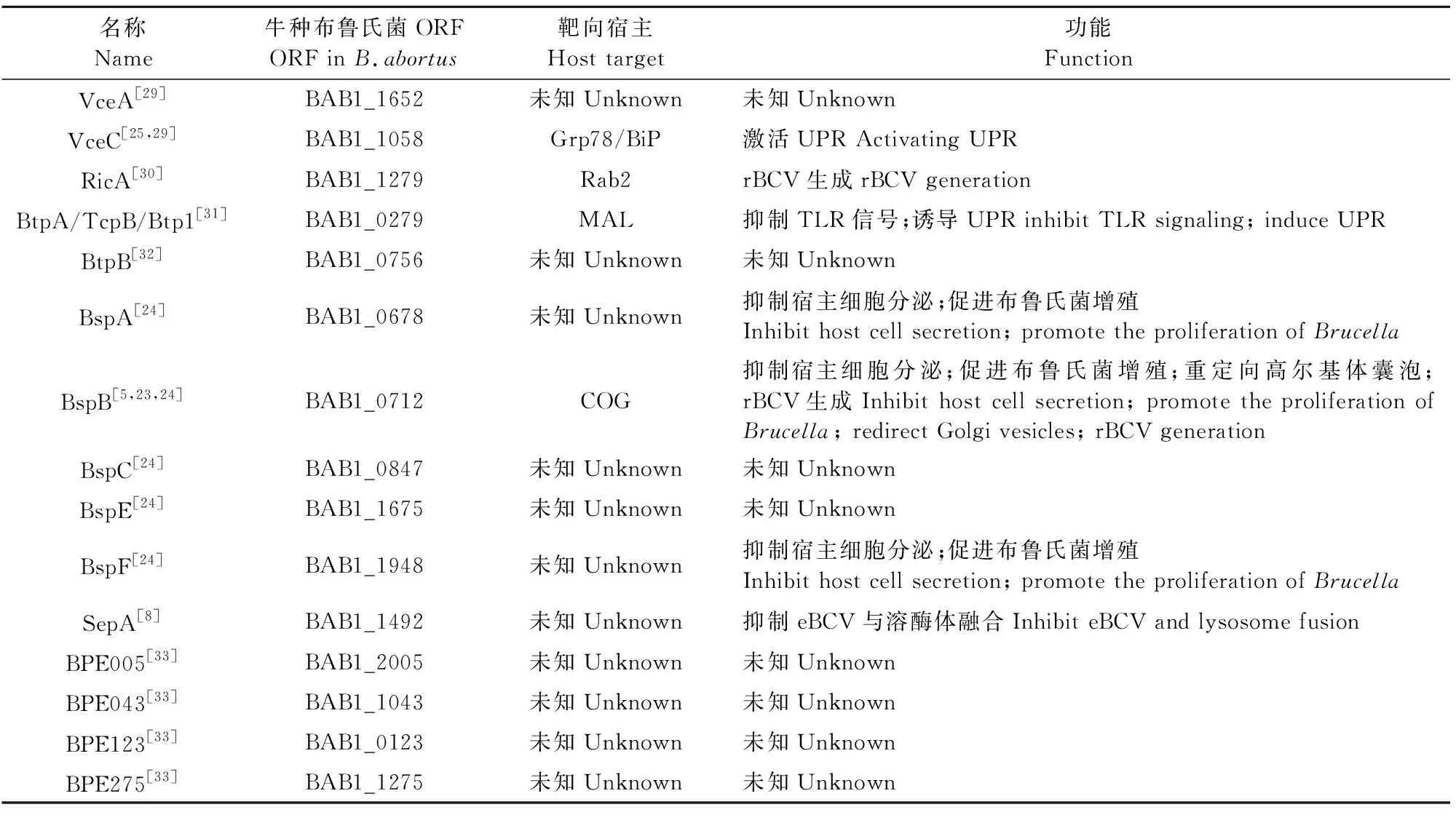
表1 布鲁氏菌T4SS效应蛋白[2,28]
T4SS效应蛋白BspA、BspB和BspF会抑制宿主蛋白分泌并促进布鲁氏菌增殖[23,24],这与布鲁氏菌特异性靶向宿主细胞多种分泌途径的假说相一致,可能是rBCV生成所必需的。虽然尚不清楚BspA和BspF的作用方式,但BspB的作用方式已被阐明[5]。BspB是rBCV生成和巨噬细胞中布鲁氏菌增殖所必需的[23]。该效应蛋白被布鲁氏菌分泌到宿主细胞中并转移至高尔基体,在这里它与保守寡聚高尔基复合体(conserved oligomeric Golgi,COG)[23]相互作用,COG是一种CATCHR家族多亚基复合体,作为高尔基体上分泌性Rab GTPases、高尔基系绳(Golgi tethers)和SNARE膜的相互作用中心,可沿着分泌途径调节高尔基体和反向囊泡分泌性转运[34]。COG的功能对于rBCV的生成和布鲁氏菌复制很重要[23],这暗示了布鲁氏菌细胞内循环与高尔基体功能具有相关性。BspB-COG的相互作用改变了COG功能,并导致依赖于COG的高尔基体反向囊泡重定向至BCV,从而使BCV获得了高尔基体来源的膜[23],表明布鲁氏菌在rBCV生成过程中可能从分泌物中吸收ER来源的膜(图2),但这一机制还有待确定。有趣的是,通过衰减Rab2的表达抑制高尔基体到ER的逆向转运过程可减弱ΔbspB缺失突变体引起的布鲁氏菌复制缺陷[23],这表明BspB可能会影响宿主细胞逆向的分泌性转运,从而将COG依赖性的高尔基体囊泡重定向到BCV。更有趣的是,T4SS效应子RicA可控制rBCV的生成过程,其可结合Rab2的GDP结合域[30],但其作用方式尚不清楚,因为它没有表现出任何GEF(鸟嘌呤核苷酸交换因子)活性,这表明布鲁氏菌分泌的几种效应蛋白可以协同作用来调节Rab2依赖性的囊泡分泌转运并促进rBCV的生成。
总之,通过研究靶向宿主分泌途径的VirB T4SS效应蛋白揭示了布鲁氏菌与宿主细胞内区室相互作用的一些机制,研究其效应蛋白的特性及破坏宿主细胞分泌途径的机制是全面了解布鲁氏菌的关键。
3.3 aBCV阶段
已知在ER结合的rBCV中增殖是布鲁氏菌致病过程的关键步骤,但数十年来,布鲁氏菌如何在增殖阶段后完成其细胞内循环仍是未知的。许多病原体致宿主细胞死亡后被释放,而布鲁氏菌却会维持其细胞内生存环境,阻止宿主细胞的死亡。Starr等[10]观察到在rBCVs增殖阶段,即感染48 h之后,布鲁氏菌藏匿位置由ER来源的细胞器转变为具有晚期内体和溶酶体特性的大囊泡,该过程伴随着LAMP1的积累和酸化。观察其超微结构发现,这些囊泡源自新月形膜结构融合吞噬体形成的rBCV,被多个膜所包围,缺乏典型的自噬标记物LC3[10]。这些囊泡被称为aBCV,其形成需要宿主细胞自噬成核功能,而不需要延伸复合物,因为beclin1、ULK1和Atg14L的缺失或沉默抑制了aBCV的形成,而自噬蛋白Atg5、Atg7、Atg4或Atg16L的缺失或沉默却没有[10]。因此,aBCV的形成应该需要自噬相关分子机制,这可能代表了另一种自噬过程或表明布鲁氏菌正在积极利用自噬途径的离散功能来产生aBCV。
重要的是,aBCV的形成与布鲁氏菌的释放紧密相关,因为aBCV形成阶段会产生新细胞的感染,而抑制aBCV的形成时则感染减少[10]。布鲁氏菌是以自由状态还是包含在结合膜的小泡中释放仍然有待确定,但最初感染的细胞释放布鲁氏菌表明其释放是一种非溶血性的胞外过程(图1)。基于aBCV的自噬特性,有人提出一个科学假说,认为布鲁氏菌的aBCV依赖性释放过程是其利用宿主细胞自噬途径的分泌功能将aBCV中的布鲁氏菌传递到细胞外环境。
在细胞内循环的最终阶段,布鲁氏菌可能通过VirB T4SS来调控aBCV的形成。rBCV的形成依赖于布鲁氏菌T4SS的表达,要证明这一说法具有一定的难度,因为无法用反向遗传学和构建VirB T4SS缺失突变体等常用方法的形式进行验证。Smith EP[35]通过控制启动子表达使 VirB11 ATPase短暂地在细胞内分泌,从而使T4SS增能,进而使布鲁氏菌生成rBCV和增殖,然后在宿主细胞内对布鲁氏菌T4SS进行灭活,结果显示,在aBCV形成和布鲁氏菌释放过程中需要VirB系统功能的表达,这表明T4SS效应子可能调控布鲁氏菌细胞内循环过程的最后阶段。未来对于T4SS具体的特性研究可能会阐明aBCV形成和布鲁氏菌释放的分子机制。
4 展望
布鲁氏菌与真核宿主紧密相关,其与宿主哺乳动物的长期共进化已将其复杂的细胞内循环过程表现为对内吞、分泌和自噬途径中细胞器室的连续利用。布鲁氏菌使用一系列T4SS效应子和其他毒力因子来调节这些区室的功能,将其原始吞噬体装配成ER相关的细胞器,随后依赖宿主细胞自噬功能以完成自身的细胞内循环,从而确保其胞内存活、增殖和释放。尽管已经确定了十几种布鲁氏菌T4SS效应子,但它们的作用方式以及对布鲁氏菌细胞内循环的作用还不甚明了。对于这些分泌蛋白功能特性的后续研究将会解决布鲁氏菌在胞内生存、致病过程以及调控宿主细胞功能等多方面问题。
