体外模拟胃肠消化及碱性蛋白酶处理后蛋清肽抗氧化活性差异及肽序列解析
2020-11-27马思彤刘静波孙惠炎魏以恒王寒颖雷洪辉刘博群
马思彤,刘静波,张 婷,王 莹,孙惠炎,魏以恒,王寒颖,雷洪辉,刘博群,*
(1.吉林省营养与功能食品重点实验室,吉林 长春 130062;2.吉林大学食品科学与工程学院,吉林 长春 130062;3.吉林工程技术师范学院食品工程学院,吉林 长春 130052)
我国是世界上鸡蛋的生产和消费大国,蛋品产业在国民经济中占有重要地位[1]。尽管中国的禽蛋产业在不断发展,但蛋品行业仍然存在诸多问题,例如,产品形式还是以传统壳蛋为主,深加工程度低,导致我国蛋品产业附加值不高,经济效益低。近几年来,诸多研究成果表明功能蛋白及蛋白肽往往具有极其重要的生物活性,尤其是功能蛋白水解后得到的活性肽片段,对机体功能环境有重要作用。
研究发现,人体内氧自由基的产生与消除失去平衡时,多余的氧自由基会导致细胞损伤,进而引起衰老、癌症等疾病[2-3]。已有大量研究表明,通过酶解得到的多肽具有很好的抗氧化活性,能有效清除体内多余自由基[4],且与传统的化学合成物作为主要来源的抗氧化剂相比,天然来源的抗氧化肽具有较好的抗氧化活性且对人体无害。因此,天然多肽将会成为越来越多消费者的选择[5]。
蛋清蛋白作为优质的蛋白质来源,通过口服的方式进入人体,在胃和肠道中进行转化。有研究表明,胃和肠道中的蛋白酶、pH值等均会影响蛋白质在人体胃肠道中的吸收与释放[6]。佟晓红等[7]研究发现体外模拟消化能够提升大豆蛋白酶解物的抗氧化活性。赵楠[8]也发现体外模拟消化能一定程度上提升松仁蛋白的抗氧化活性。虽然人体自身的蛋白酶能够一定程度上水解蛋白质,使其发挥生物活性。但即使是同一蛋白质,在不同的酶解方式、pH值或无机盐环境中,都会水解出不同种类的肽段[9]。因此,本研究通过对比碱性蛋白酶酶解及体外模拟胃肠消化2 种方式处理蛋清蛋白后水解物的抗氧化活性差异,明确蛋白质深加工对于蛋清肽抗氧化活性的影响,以期为蛋品深加工的方法和意义提供一定的实验依据。
1 材料与方法
1.1 材料与试剂
蛋清蛋白粉 吉林金翼蛋品有限公司;HepG2细胞 中国科学院上海生命科学研究院细胞库;DMEM高糖培养基、胎牛血清、磷酸盐缓冲液(phosphate buffered saline,PBS)、胰蛋白酶-EDTA 美国Gibco公司;碱性蛋白酶Alcalase 丹麦诺维信生物技术有限公司;胃蛋白酶、胰液 美国Genview Scientific公司;乙腈、甲酸(色谱纯) 美国Thermo Fisher Scientific公司;水溶性VE(Trolox)、2,2’-联氮-双-3-乙基苯并噻唑啉-6-磺酸(2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid),ABTS)、2,2-偶氮二(2-甲基丙基咪)二盐酸盐(2,2’-azobis(2-methyl-propanimidamide)dihydrochloride,AAPH) 美国Sigma公司;MTS单细胞溶液增殖检测试剂盒 美国Promega公司;细胞内活性氧(reactive oxygen species,ROS)检测试剂盒上海碧云天生物技术有限公司;H2O2、荧光素钠 天津化学有限公司。
1.2 仪器与设备
多功能酶标仪 美国伯腾仪器有限公司;膜分离装置 美国Millipore公司;EASY-nLC 1000超高效液相色谱-串联质谱(ultra performance liquid chromatography/tandem mass spectrometry,UPLC-MS/MS) 美国Thermo公司;细胞培养箱 上海力申科学仪器有限公司;移液枪 德国Eppendorf公司;培养皿、离心管及96 孔板 美国Corning公司。
1.3 方法
1.3.1 蛋清肽的制备
1.3.1.1 碱性蛋白酶酶解
参照Wang Liying等[10]的方法并稍作修改。称取100 g蛋清蛋白粉,加入2 L蒸馏水配制成质量分数5%的蛋清蛋白溶液。将溶液在90 ℃下水浴10 min,使蛋白质发生变性裂解,然后降温至50 ℃。用1 mol/L NaOH溶液将体系的pH值调至10,加入质量分数4%的碱性蛋白酶Alcalase,保持温度50 ℃、pH 10持续酶解3 h。之后将体系沸水浴10 min灭活蛋白酶,冷却至室温,用1 mol/L的HCl溶液将体系的pH值调整为7(后续将用来处理细胞)。将酶解液离心,在4 ℃下10 000 r/min离心10 min,收集上清液,-20 ℃贮存备用。
1.3.1.2 体外模拟胃肠消化
参照Alting等[11]报道的方法并稍作修改。称取20 g蛋清蛋白粉,加入2 L蒸馏水配制成质量分数1%的蛋清蛋白溶液。按1∶50(m/m)加入胃蛋白酶((EC 3.4.23.1),来源于猪胃黏膜,酶活力为3 000 U/mg),用1 mol/L的HCl溶液将pH值调至2,保持体系温度在37 ℃,持续酶解90 min。用1 mol/L的NaOH溶液调pH值至7,按1∶25(m/m)加入胰液((EC 3.4.21.4),来源于猪胰腺,酶活力为250 U/mg),保持温度3 7 ℃和p H 7,持续酶解4 h。将体系沸水浴10 min灭活蛋白酶,冷却至室温。将酶解液在4 ℃下10 000 r/min离心10 min,收集上清液,-20 ℃贮存备用。
1.3.2 蛋清肽的分离纯化
将酶解上清液用截留分子质量为1、3 kDa和10 kDa的超滤膜进行分离,收集分子质量小于1 kDa、1~3 kDa和3~10 kDa的蛋清肽组分,真空冷冻干燥后置于-20 ℃干燥保存备用。
1.3.3 蛋清肽的ABTS阳离子自由基清除能力测定
取ABTS 10 mg溶于2.6 mL蒸馏水中配制为7 mmol/L的ABTS溶液,取3.45 mg过硫酸钾溶于2.6 mL蒸馏水中配制成2.45 mmol/L的过硫酸钾溶液,将两者按体积比1∶1混合配制为ABTS储备液,使用前在37 ℃下避光静置12 h后,用0.2 mol/L pH 7.4的PBS稀释ABTS储备液(20~30 倍),使其在734 nm波长处的吸光度为0.70±0.02,配制为ABTS工作液,现用现配。实验分组如下:1)空白组:每孔加入200 μL PBS;2)对照组:每孔加入180 μL ABTS工作液和20 μL PBS;3)实验组:每孔加入180 μL ABTS工作液和20 μL样品溶液(终质量浓度分别为0.50、1.00、5.00、10.00 μg/mL),设置3 个复孔。在96 孔板中,按照上述实验分组加入ABTS工作液、样品溶液以及PBS,振荡10 s混匀后在37 ℃下静置反应5 min,在734 nm波长处测定反应体系的吸光度。ABTS阳离子自由基清除率按式(1)计算。
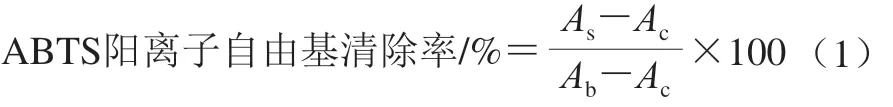
式中:As、Ab、Ac分别为实验组、空白组、对照组的吸光度。
1.3.4 蛋清肽的氧自由基吸收能力测定
氧自由基吸收能力(oxygen radical absorbance capacity,ORAC)测定参考Zheng Lin等[12]的方法并略作修改。以75 mmol/L PBS(pH 7.4)为溶剂,分别配制荧光素钠溶液(终浓度为70 nmol/L)、AAPH溶液(终浓度为12 mmol/L)和样品溶液(终质量浓度为10 μg/mL),Trolox溶液设置为阳性对照(终质量浓度为10 μg/mL)。实验分组如下:1)空白组:每孔加入120 μL荧光素钠溶液和20 μL PBS;2)阳性对照组:每孔加入120 μL荧光素钠溶液和20 μL Trolox溶液;3)实验组:每孔加入120 μL荧光素钠溶液和20 μL样品溶液,设置3 个复孔。在黑色96 孔板中,按照上述实验分组加入荧光素钠溶液、样品溶液、Trolox溶液以及PBS,振荡10 s混匀后在37 ℃下静置预热2 min,然后每孔中快速加入60 μL AAPH溶液启动反应,立即将黑色96 孔板放入酶标仪中。反应体系在37 ℃下检测,相邻两个测定点之间的时间间隔设置为2 min,每次读数前振板2 s,连续测定180 min(荧光衰减趋于稳定为止)。设置酶标仪的激发波长为485 nm,发射波长为520 nm。样品的抗氧化能力与AAPH自由基作用下荧光衰退曲线的延缓部分面积直接相关。
1.3.5 细胞培养
HepG2细胞培养基配方为:88%(体积分数,下同)DMEM高糖培养基、10%胎牛血清、1%非必需氨基酸和1%双抗。培养箱内环境保持在37 ℃和5% CO2,细胞每周传代2~3 次,日常视情况更换培养基。待细胞呈对数生长时进行后续实验。
1.3.6 HepG2细胞氧化损伤模型的建立
细胞存活率的检测采用单溶液细胞增殖检测试剂盒,即MTS法[13]。
在96 孔板中接种HepG2细胞,密度为1.0×104cells/mL,每孔加入80 μL细胞悬液。在培养箱中培养12 h后,每孔中加入10 μL DMEM培养基孵育12 h。再按照实验分组分别加入H2O2和DMEM培养基孵育6 h后,每孔加入20 μL MTS孵育1~4 h,在490 nm波长处测定96 孔板的吸光度。
实验分组设置如下:1)空白组:每孔加入100 μL DMEM培养基;2)对照组:每孔加入80 μL细胞悬液和20 μL DMEM培养基;3)实验组:每孔加入80 μL细胞悬液、10 μL DMEM培养基和10 μL H2O2溶液(终浓度为200、300、400、500、600、700 μmol/L),设置6 个复孔。细胞相对存活率按式(2)计算。

式中:As、Ab、Ac分别为实验组、空白组、对照组的吸光度。
1.3.7 蛋清肽对H2O2诱导氧化损伤的HepG2细胞的保护作用
在96 孔板中接种HepG2细胞,密度为1.0×104cells/mL,每孔加入80 μL细胞悬液。在培养箱中培养12 h后,每孔中加入10 μL DMEM培养基或者蛋清肽溶液孵育12 h。再按照实验分组分别加入DMEM培养基或者H2O2孵育6 h后,每孔加入20 μL MTS孵育1~4 h,在490 nm波长处测定96 孔板样品的吸光度。
实验分组设置如下:1)空白组:每孔加入100 μL DMEM培养基;2)对照组:每孔加入80 μL细胞悬液和20 μL DMEM培养基;3)损伤组:每孔加入80 μL细胞悬液、10 μL DMEM培养基和10 μL H2O2溶液(终浓度为500 μmol/L);4)阳性对照组:每孔加入80 μL细胞悬液、10 μL蛋清肽溶液(终质量浓度分别为0.01、0.1、1 mg/mL)和10 μL DMEM培养基;5)实验组:每孔加入80 μL细胞悬液、10 μL蛋清肽溶液(终质量浓度分别为0.01、0.1、1 mg/mL)和10 μL H2O2溶液(终浓度为500 μmol/L),设置6 个复孔。
1.3.8 细胞内ROS水平的检测
按试剂盒说明书进行ROS水平检测。
1.3.9 蛋清源多肽的氨基酸序列鉴定
1.3.9.1 样品前处理蛋清肽通过葡聚糖凝胶色谱柱(G15)脱盐后冷冻干燥,用于UPLC-MS/MS分析。
1.3.9.2 色谱条件
流动相A:0.1%甲酸溶于2%乙腈;流动相B:0.1%甲酸溶于98%乙腈。洗脱程序:0~16 min,流动相B 5%~23%;16~22 min,流动相B 23%~35%;22~27 min,流动相B 35%~80%;27~30 min,流动相B保持80%。流速:0.4 mL/min;进样量:10 μL。
1.3.9.3 质谱条件
离子源:电喷雾离子化;扫描模式:正离子模式;裂解电压:120 V;碰撞电压:20 V;质量扫描范围m/z 50~1 000。
1.3.10 纯肽合成
通过高效液相色谱-串联质谱解析出混合多肽的序列,以此为依据通过固相合成制备纯肽,纯肽合成工作委托生工生物工程(上海)股份有限公司进行,纯度达到98%以上。
1.3.11 纯肽的ABTS阳离子自由基清除能力测定
方法同1.3.3节。
1.3.12 纯肽的ORAC测定
方法同1.3.4节。
1.4 数据统计分析
2 结果与分析
2.1 蛋清肽ABTS阳离子自由基清除能力的变化
ABTS阳离子自由基清除能力的测定因其快捷、高效、可信度高,被广泛应用于评价水溶性物质的抗氧化活性[14]。如图1所示,未经处理的蛋清蛋白ABTS阳离子自由基清除能力很差,在经过碱性蛋白酶酶解或者体外模拟胃肠消化后的蛋清蛋白被分解为多肽或者氨基酸,活性基团得以暴露;因此,在同一质量浓度条件下,和未经处理的蛋清蛋白相比,处理后的蛋清肽ABTS阳离子自由基清除能力都得到提高,其中分子质量小于1 kDa的蛋清肽抗氧化活性高于其他分子质量的组分,随着分子质量的增大,蛋清肽的抗氧化活性减弱。这一结果与迟玉杰等[15]的研究一致,其研究发现,蛋清肽分子质量越小,其抗氧化活性越强。同时,蛋清肽质量浓度越高,抗氧化活性越强,这也与图1结果相对应。除此之外,在同一质量浓度下,不同处理方式得到的蛋清肽抗氧化能力也有显著差异。实验发现,碱性蛋白酶酶解物的抗氧化活性普遍优于体外模拟胃肠消化产物,其中分子质量小于1 kDa的碱性蛋白酶酶解物的抗氧化活性显著高于其他组分(P<0.05)。这可能是由于碱性蛋白酶酶解处理后的蛋清肽暴露出的活性位点更多,蛋清肽可能作为氢供体、自由基稳定剂等方式来抑制过氧化作用。占今舜等[16]用不同酶解方法处理的蛋清肽改善小鼠血清生化指标和肝脏抗氧化能力,并发现Alcalase水解物效果最好。Zheng Zhaojun等[17]研究发现黑豆的菠萝蛋白酶和碱性蛋白酶酶解产物抗氧化活性好于其他酶酶解产物。

图1 蛋清肽的ABTS阳离子自由基清除能力Fig. 1 ABTS radical cation scavenging properties of egg white peptides
2.2 蛋清肽氧自由基吸收能力的变化
由于氧自由基吸收能力可以直接反映抗氧化剂阻断自由基链式反应的能力,被认为是最接近人体生理系统真实反应情况的实验方法[18]。ORAC反应是典型的基于氢原子转移机制的氧化过程,氢原子转移是自由基链式反应中的关键步骤,反应后会形成更加稳定的自由基[19]。

图2 蛋清肽的氧自由基吸收能力Fig. 2 Oxygen radical absorption capacity of egg white peptides
图2 表示蛋清肽在质量浓度为10 μg/mL时的荧光衰退曲线,在对照组(PBS代替抗氧化剂)中,荧光衰减最快,实验组荧光衰减较对照组均放缓,说明蛋清肽都有抗氧化活性。其中,分子质量小于1 kDa碱性蛋白酶酶解物抗氧化活性最强,同ABTS阳离子自由基清除能力的结果一致。Trolox属于VE的水溶性类似物,是一种常用的抗氧化剂和自由基清除剂,在这里被用作阳性对照,可以看出和Trolox相比,蛋清肽的抗氧化活性仍存在一定差距。
分别计算蛋清肽的ORAC,分子质量小于1 kDa碱性蛋白酶酶解物的ORAC最高,为(0.399±0.180)μg/mL,其次是分子质量小于1 kDa体外模拟胃肠消化产物((0.34±0.05)μg/mL),分子质量1~3 kDa的碱性蛋白酶酶解物和体外模拟胃肠消化产物的ORAC分别为(0.31±0.13)μg/mL和(0.21±0.11)μg/mL,未经处理的蛋清蛋白与3~10 kDa体外模拟胃肠消化产物活性接近。
2.3 HepG2细胞氧化损伤模型的建立
H2O2可诱导细胞氧化损伤,刺激细胞内ROS的生成,打破细胞内氧化还原反应平衡,从而增强细胞氧化应激,最终可导致细胞损伤和凋亡[20]。
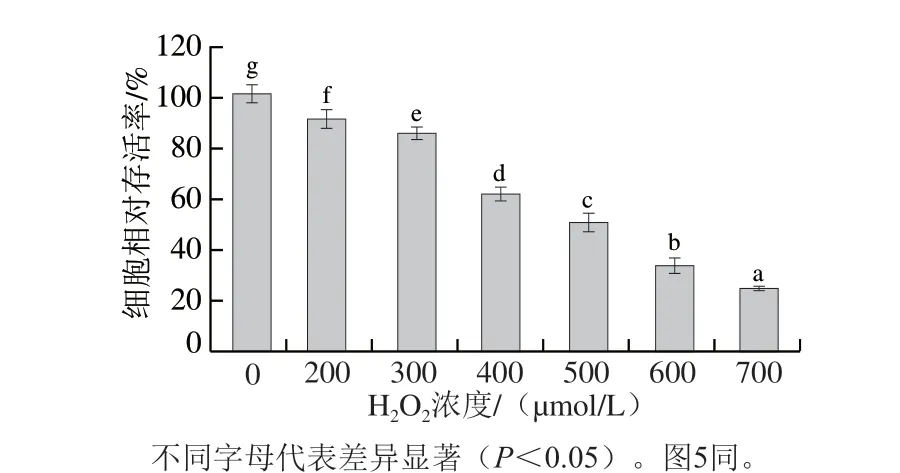
图3 不同浓度H2O2对HepG2细胞相对存活率的影响Fig. 3 Effect of H2O2 concentration on survival rate of HepG2 cells
由图3可知,随H2O2浓度的升高,HepG2细胞的相对存活率降低,即氧化损伤作用越强。当H2O2的终浓度为500 μmol/L时HepG2细胞的相对存活率为(49.96±0.04)%,不同H2O2浓度之间的细胞存活率均有显著差异(P<0.05),因此选择500 μmol/L的H2O2建立HepG2细胞氧化损伤模型,用于后续实验。
2.4 蛋清肽对HepG2细胞氧化损伤的影响
如图4所示,在500 μmol/L的H2O2作用下,分子质量小于1 kDa的碱性蛋白酶酶解物活性最高,几乎可以保护细胞免受氧化应激损伤,中质量浓度组及高质量浓度组和对照组相比无显著差异(P>0.05),细胞相对存活率分别为(96.62±0.05)%和(102.87±0.08)%。分子质量3~10 kDa的体外模拟胃肠消化产物活性最弱,与损伤组相比各质量浓度均无显著差异(P>0.05),这可能是由于该组分蛋清肽分子质量较大,难以被细胞吸收利用。其他组分蛋清肽和损伤组相比均有显著差异,说明不同组分蛋清肽均能不同程度上对细胞氧化损伤有保护作用,且碱性蛋白酶酶解物的活性优于体外模拟胃肠消化产物。可能是因为碱性蛋白酶酶解物能够一定程度上弥补人体无法通过自身消化产生的肽段,且肽段种类更多,从而在细胞内有更高的活性。除了处理方式对蛋清肽活性有影响外,蛋清肽质量浓度也会影响蛋清肽活性,蛋清肽质量浓度越高,蛋清肽的活性越强。He Ying等也曾发现鸭胚胎肽的碱性蛋白酶酶解物能够保护HepG2细胞免受H2O2诱导的氧化应激损伤[21]。
为排除蛋清肽毒性作用的影响,在未加H2O2损伤的情况下,检测最高质量浓度(1 mg/mL)蛋清肽对HepG2细胞生长的影响,实验组和对照组细胞相比存活率均无显著性差异(P>0.05),说明蛋清肽本身对HepG2细胞无毒副作用。排除高质量浓度蛋清肽刺激细胞的可能性后,蛋清肽对HepG2细胞氧化损伤具有一定的保护作用,可能是通过抑制氧化应激实现的。
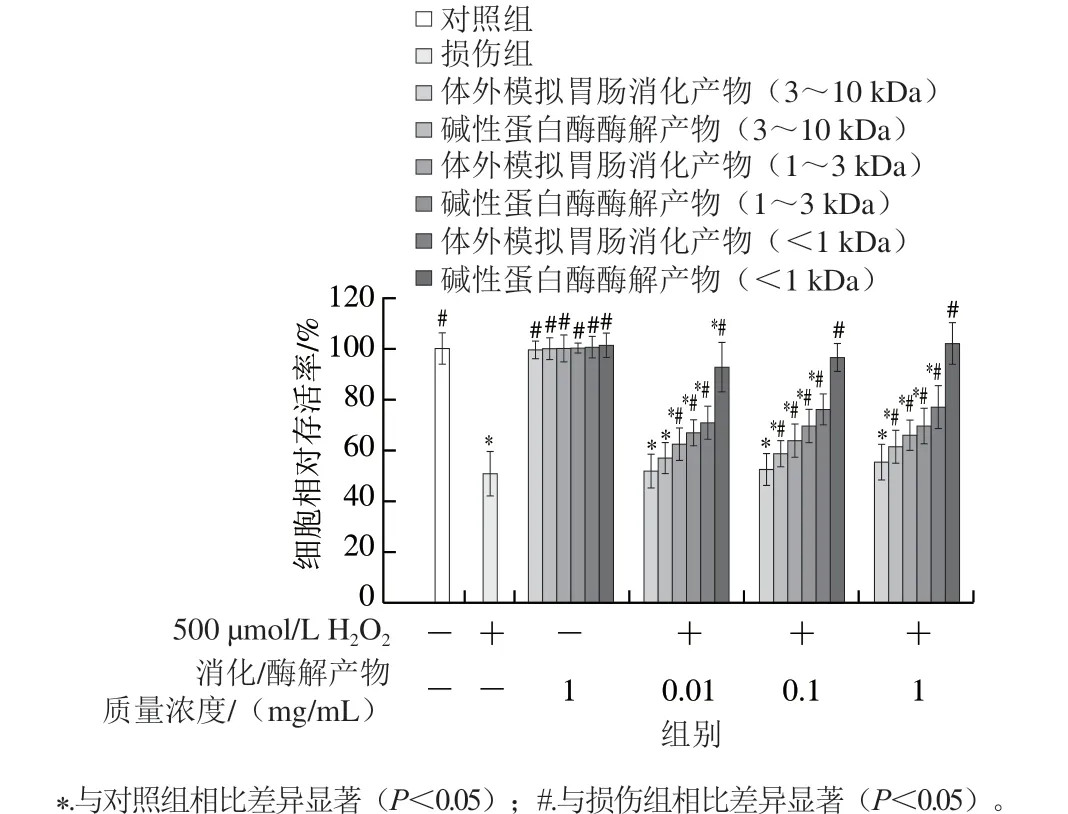
图4 蛋清肽对HepG2细胞相对存活率的影响Fig. 4 Effect of egg white peptides on survival rate of HepG2 cells exposed to H2O2
2.5 蛋清肽的细胞内ROS清除活性
二 氯 二 氢 荧 光 素 - 乙 酰 乙 酸 酯(2’,7’-dichlorofluorescin diacetate,DCFH-DA)是一种自身没有荧光且可以自由穿透细胞膜的荧光染料。细胞内ROS将非荧光的DCFH-DA氧化为荧光的2’,7’-二氯荧光素(2’,7’-dichlorofluorescein,DCF)。通过检测DCF的荧光强度,可以得到细胞内ROS的水平,即DCF荧光强度越强,细胞内ROS水平越高[22]。如图5所示,H2O2处理后,HepG2细胞内DCF荧光强度显著增强(P<0.05),大约是对照组的3 倍,表明H2O2诱导HepG2细胞内产生大量ROS。经过蛋清肽预处理的HepG2细胞内ROS水平有不同程度的减少,且呈浓度依赖性。其中分子质量小于1 kDa的蛋清肽和损伤组相比细胞内ROS水平显著降低(P<0.05),尤其是分子质量小于1 kDa的碱性蛋白酶酶解物,即使在低质量浓度(0.01 mg/mL)下也能很好地抑制ROS对HepG2细胞的损伤,该组分的DCF荧光强度和对照组相比无显著差异,好于体外模拟胃肠消化产物活性。Yarnpakdee[23]和Zha Fengchao[24]等的研究显示,从虾和罗非鱼中提取的抗氧化肽也具有清除HepG2细胞内ROS的作用。
图5 蛋清肽对H2O2诱导的氧化损伤HepG2细胞内ROS水平的影响Fig. 5 Effect of egg white peptides on intracellular ROS in HepG2 cells with H2O2-induced oxidative damage
2.6 UPLC-MS/MS鉴定蛋清肽的序列
分析对比不同处理方式得到的蛋清肽抗氧化活性后,发现分子质量小于1 kDa的碱性蛋白酶酶解产物具有较高的抗氧化活性,因此利用质谱方法分析该组分蛋清肽的序列,用于后续研究。为了得到蛋清源抗氧化肽的氨基酸序列,根据Zhang Binying等[25]的报道,使用从头测序的方法解析未知多肽的二级MS碎片信息,从而确定多肽的氨基酸序列。多肽一般在电喷雾离子源条件下发生质子化,当碰撞能量低于200 eV时,多肽大多在酰胺键位置断裂,形成b离子和y离子,因此,MS/MS碎片离子主要为b离子和y离子[26]。


图6 蛋清肽鉴定的二级质谱图Fig. 6 Tandem mass spectra of identified egg white peptides
如图6所示,解析分子质量小于1 kDa的碱性蛋白酶酶解产物得到四肽FYCP、YCPI、YLPR。这3 条多肽都是源于蛋清蛋白中含量最高的卵白蛋白,其中YLPR曾被Ayako等[27]证明具有减缓小鼠焦虑的活性。
2.7 蛋清源活性肽的ABTS阳离子自由基清除能力
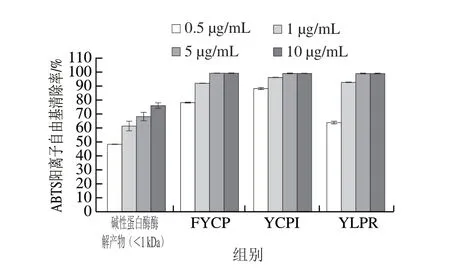
图7 蛋清源活性肽的ABTS阳离子自由基清除能力Fig. 7 ABTS radical cation scavenging properties of bioactive peptides from egg white proteins
为验证从分子质量小于1 kDa的碱性蛋白酶酶解物中解析出的3 条多肽FYCP、YCPI、YLPR的抗氧化活性,对这3 条肽的ABTS阳离子自由基清除能力进行测定,并与分子质量小于1 kDa的碱性蛋白酶酶解产物进行比较。如图7所示,3 条多肽的抗氧化活性都明显优于分子质量小于1 kDa的碱性蛋白酶酶解产物,当多肽质量浓度达到5 μg/mL时就能完全清除ABTS阳离子自由基,且FYCP和YCPI的抗氧化活性不相上下。这3 条多肽中均含有Tyr和Pro,这类氨基酸的吲哚基和苯环可供氢,因此具有良好的ABTS阳离子自由基清除能力。Hernández-Ledesma等[28]从发酵乳中提取抗氧化肽,并得出类似的结论。
2.8 蛋清源活性肽的氧自由基吸收能力
图8显示了3 种多肽和分子质量小于1 kDa的碱性蛋白酶酶解物氧自由基吸收能力测定的荧光衰退曲线,可以看到FYCP的活性最强,ORAC为(2.42±0.13)μg/mL,其次是YCPI,ORAC为(2.04±0.19)μg/mL。这两条肽的抗氧化活性都要高于阳性对照Trolox,远远高于分子质量小于1 kDa的碱性蛋白酶酶解物((0.399±0.180)μg/mL)。但与ABTS阳离子自由基清除能力的结果不同,YLPR的氧自由基吸收能力(ORAC为(0.24±0.16)μg/mL)弱于分子质量小于1 kDa的碱性蛋白酶酶解物。这种差异可能是由于两种测定方法的作用机制不同,ORAC法是基于氢原子转移机制[29];ABTS阳离子自由基清除能力的测定是根据被氧化的ABTS变为蓝绿色的ABTS阳离子,当被测样品具有抗氧化活性时,颜色减褪。此外,Zhang Deju等[30]的研究中显示,Cys在20 种氨基酸中抗氧化活性最高,在氧自由基吸收能力中发挥重要作用。3 种多肽同样含有Tyr和Pro,但YLPR不含Cys,这可能是YLPR的氧自由基吸收能力小于FYCP和YCPI的原因。
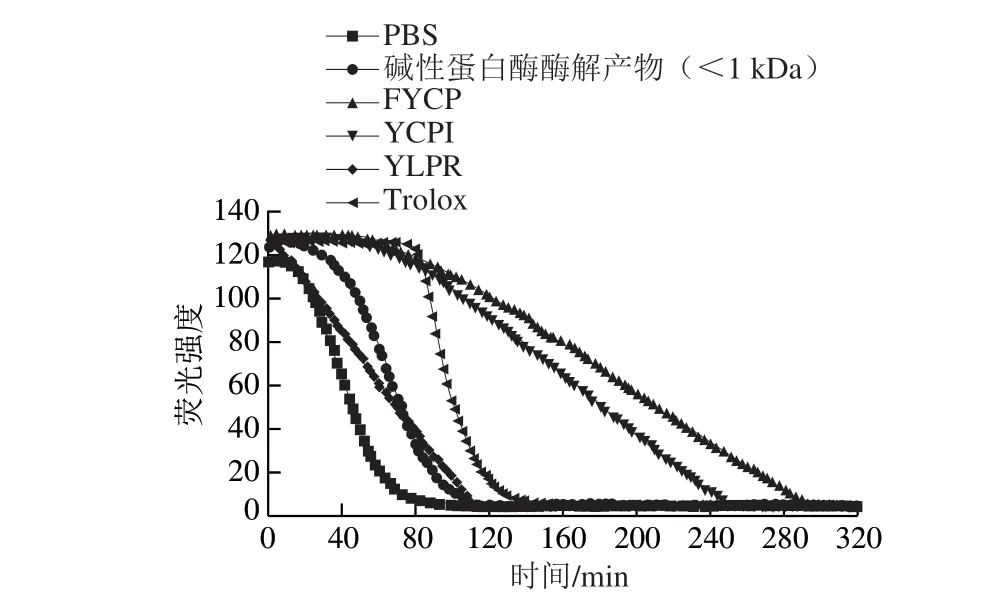
图8 蛋清源活性肽的氧自由基吸收能力Fig. 8 Oxygen radical absorption capacity of bioactive peptides from egg white proteins
3 结 论
通过对比碱性蛋白酶及体外模拟胃肠消化处理后蛋清肽抗氧化活性差异,碱性蛋白酶酶解物的抗氧化活性最强,其次是体外模拟胃肠消化产物,未经处理的蛋清蛋白抗氧化活性十分微弱,这表明,对蛋清蛋白进行深加工可能有更高的生物活性和经济价值。此外,蛋清肽的抗氧化活性还与其分子质量和浓度密切相关,小分子质量高浓度的蛋清肽具有更强的抗氧化活性。其中分子质量小于1 kDa的碱性蛋白酶解物能够有效抑制H2O2诱导的HepG2细胞内ROS的产生,进而阻止细胞的氧化应激损伤和凋亡。利用UPLC-MS/MS从该组蛋清源活性肽中解析出3 条四肽FYCP、YCPI、YLPR,均具有良好的抗氧化活性。以上研究结果在一定程度上说明了蛋白质深加工的重要性,并为蛋清源生物活性肽的抗氧化活性研究提供了一定的参考。
