牦牛ATG5多克隆抗体的制备及应用
2023-12-08潘阳阳焦正兴王靖雷马文斌王军乾余四九王立斌
高 翔,潘阳阳,2,王 萌,2,焦正兴,王靖雷,马文斌,王军乾,余四九,2,崔 燕,2,王立斌,2*
(1.甘肃农业大学 动物医学院,甘肃 兰州 730070;2.甘肃省牛羊胚胎工程技术研究中心,甘肃 兰州 730070)
自噬(autophagy)是真核细胞中普遍存在的生命现象,是将细胞内变形、衰老或损伤的蛋白质和细胞器转运到溶酶体腔中消化降解的一种代谢过程,在维持细胞正常功能中起着重要作用[1-2]。哺乳动物卵泡发育过程中,自噬有助于维持健康的原始卵泡数量、生殖细胞存活和去除黄体残余物并诱导细胞凋亡[3-4],对卵巢闭锁和生殖细胞发育进行调控[5-6]。自噬相关蛋白通过介导多种信号诱导自噬以维持雄性生殖细胞的发育过程[7]。当自噬相关基因在雄性生殖细胞中被破坏后,将导致精子顶体发育异常、线粒体重排以及多余细胞质残留以及受精卵个体发育失败[8]。研究表明,子宫内膜在卵巢激素的调控下发生自噬,导致免疫细胞浸润,对子宫内膜脱落、组织修复和预防感染起着不可或缺的作用[9]。在胚胎附植过程中,子宫内膜自噬水平也发生显著变化,附植前期子宫内膜细胞中自噬水平较高,随着附植完成,自噬水平出现显著下调[10],说明自噬在维持妊娠中发挥重要作用。
在已知的自噬相关 (autophagy-related,ATG)蛋白中,ATG5蛋白是自噬囊泡形成不可或缺的[11]。ATG5蛋白作为自噬与细胞凋亡之间的分子开关,它的缺失可导致细胞自噬水平下调或完全被抑制[12]。在自噬体囊泡伸长和成熟过程中,ATG7蛋白和ATG10蛋白促进ATG12蛋白和ATG5蛋白结合形成二聚体复合物 (ATG12-ATG5-ATG16L1),这是微管相关蛋白1轻链3(microtubule-associated protein 1 light chain 3,LC3)和磷脂酰乙醇胺(phosphatidylethanolamine,PE)之间形成共价键所必需的[13]。而LC3前体在ATG4蛋白的作用下转化为LC3-Ⅰ,由ATG7蛋白激活并转移到ATG3蛋白。ATG12-ATG5-ATG16L1 复合物促进LC3-Ⅰ从ATG3蛋白转移到PE以产生LC3Ⅱ,它是自噬通量的标准标记,定位于自噬体的内膜和外膜[14-17]。
牦牛(Bosgrunniens)是生活在高海拔地区的特有畜种,常年放牧饲养,生活环境气候条件非常恶劣,高寒低氧。牦牛主要分布在我国青藏高原,占世界牦牛总数的95%,是当地居民重要的生活物资来源,有很高的经济价值。受到自然环境的影响,牦牛的生产性能和繁殖性能比较低下,且人们对其生殖生理活动的研究相对较少。本研究通过合成牦牛ATG5基因,构建重组质粒,诱导重组蛋白表达,并用表达的蛋白免疫日本大耳白兔,制备牦牛ATG5多克隆抗体,并检测ATG5蛋白在牦牛组织中的表达情况,为进一步利用辅助生殖技术提高牦牛的繁殖率提供理论依据。
1 材料与方法
1.1 实验动物与样品采集
1.1.1实验动物 日本大耳白兔购自兰州兽医研究所,10月龄,雌、雄各1/2,室内饲养,饮水充足,健康状况良好。
1.1.2牦牛组织样品采集 本研究所用的牦牛组织样品采集自青海省西宁市某屠宰场。健康成年牦牛经颈动脉放血屠宰后,采集组织样品,分别置于4%多聚甲醛溶液和液氮中用于后期试验。牦牛输卵管上皮细胞由甘肃省牛羊胚胎工程技术研究中心提供。
1.2 主要仪器与试剂
1.2.1主要仪器 超声波细胞破碎仪,宁波新芝生物公司;酶标仪,美国Thermo公司;显微拍照仪,日本Olympus公司。
1.2.2主要试剂 原核表达载体pET-32a由甘肃省牛羊胚胎工程技术研究中心保存; 5-溴-4-氯-3-吲哚-β-D-半乳糖苷(5-bromo-4-chloro-3-indolyl-beta-D-galactopyranoside,X-gal)、异丙基-β-D-硫代半乳糖苷(isopropyl-β-D-thiogalactoside,IPTG),北京Solarbio公司;亲和层析柱料和佐剂,默克(中国)有限公司;感受态细胞BL21(DE3),大连TaKaRa公司;磷酸缓冲盐溶液(phosphate-buffered saline,PBS),Gibco (美国)公司;HistostainTM-Plus Kits免疫组化染色试剂盒,博奥森(北京)公司。
1.3 牦牛pET-32a-ATG5重组质粒的构建
1.3.1人工合成目的基因 根据GenBank公布的牦牛ATG5基因序列(登录号:MK531791),由武汉戴安生物技术有限公司人工合成牦牛ATG5基因片段,长度为840 bp,包含限制性内切酶Hind Ⅲ和BamHⅠ位点。
1.3.2构建重组质粒 将获得的ATG5基因片段与pET-32a(含有His与Trx标签,蛋白相对分子质量大小为20 kDa)质粒连接,构建pET-32a-ATG5重组质粒。使用内切酶XhoⅠ和ApaBⅠ对重组质粒进行双酶切验证。
1.4 牦牛ATG5重组蛋白原核表达将pET-32a-ATG5重组质粒转化至BL21(DE3)感受态细菌,恒温箱培养过夜。挑选6个单克隆, 37℃ 220 r/min培养至菌液D600 nm=0.5~0.6时,加入终浓度0.5×10-3mol/L IPTG。20℃恒温诱导3.5 h,通过SDS-PAGE筛选表达好的菌株。将表达良好的菌种接种至200 mL抗性培养基中,37℃ 220 r/min培养过夜。加入新鲜抗性培养液至800 mL,培养2 h,至菌液D600 nm=0.5~0.6,加入终浓度1 mol/L IPTG,37℃诱导3.5 h。
1.5 牦牛ATG5重组蛋白的纯化将1.4所得菌液4℃条件下,4 000 r/min离心15 min收集菌体,弃上清。PBST重悬菌体,加入终浓度1×10-3mol/L PMSF,超声破碎6 min。220 r/min,4℃孵育1 h;8 000 r/min 离心15 min,收取上层清液。加入含有400 μL 纯化树脂的层析柱中,4℃结合过夜。2 000 r/min 离心5 min,收集纯化树脂后用0.02 mol/L 咪唑洗涤液清洗2次;加入300 μL浓度为0.3 mol/L的咪唑洗脱液,4℃孵育1 h,离心收集上清。再次加入300 μL的洗脱液,静置1 h,离心收集上清,将2次洗脱液合为1管。SDS-PAGE鉴定蛋白相对分子质量。
1.6 牦牛ATG5多克隆抗体的制备
1.6.1动物免疫及抗血清的获得 利用纯化后的ATG5重组蛋白免疫2只日本大耳白兔,首次免疫时使用弗氏完全佐剂,加强免疫使用弗氏不完全佐剂。4次加强免疫后,采集血清。置于4℃冰箱静置过夜,使血清充分析出。4℃ 4 000 r/min离心5 min得到分离的抗血清,-80℃保存备用。
1.6.2Western blot检测抗血清特异性 取ATG5重组蛋白,100℃水浴10 min 进行蛋白SDS变性。配置10%的分离胶溶液和5%的浓缩胶溶液,向样品槽中分别加入15 μL样品,进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)分离蛋白。将蛋白转移至PVDF膜,一抗为所制备的日本大耳白兔血清,稀释比例为1∶1 000,4℃孵育过夜;洗涤后,孵育山羊抗兔二抗,稀释比例为1∶8 000,室温水平摇床孵育1 h,洗涤后曝光。
1.6.3牦牛ATG5多克隆抗体的纯化 室温下,将1 mg纯化蛋白质与溴化氢活化的 Sepharose 4B柱颠倒混匀1 h,制备亲和纯化柱;10 mL抗血清与亲和纯化柱孵育过夜;pH=5.0的HCl预洗,除去杂抗体,pH=2.5,0.15 mol/L的甘氨酸电泳缓冲液洗脱,10×PBS缓冲液中和,制备亲和纯化抗体;对PBS缓冲液透析换液,-80℃ 保存备用。
1.7 牦牛ATG5多克隆抗体的效价及特异性检测
1.7.1间接ELISA法检测牦牛ATG5多克隆抗体的效价 使用碳酸盐缓冲液稀释纯化的牦牛ATG5重组蛋白,按照每孔100 ng 加入聚苯乙烯96孔板中,4℃ 孵育过夜;牦牛ATG5多克隆抗体,按1∶10 000,1∶20 000,1∶40 000,1∶80 000,1∶160 000,1∶320 000,1∶640 000,1∶1 280 000,1∶2 560 000,1∶5 120 000,1∶10 240 000进行倍比稀释(空白血清做阴性对照),每孔100 μL,温室孵育1 h,弃去孔内液体,洗涤3次。每孔加入100 μL HRP标记的山羊抗兔IgG,37℃孵育40 min,弃去孔内液体,洗涤3次。每孔加100 μL显色液,温箱暗处放置30 min,加入50 μL终止液终止显色。以D450 nm阳性血清/D450 nm阴性血清>2.1的最大稀释倍数为抗体效价。
1.7.2免疫荧光检测牦牛ATG5多克隆抗体的特异性 选取原代牦牛输卵管上皮细胞,调整细胞密度为1×105个/mL,铺于赖氨酸包备的盖玻片上,培养箱培养5 h,使细胞贴壁;PBS清洗3次,1 min/次;2%多聚甲醛固定1 h;PBS清洗3次,5 min/次;0.5% Triton X-100透化20 min;PBS清洗3次,5 min/次;1% BSA封闭2 h;PBS清洗3次,5 min/次;牦牛ATG5多克隆抗体(1∶500)4℃孵育过夜;PBS 清洗3次,5 min/次;加入FITC标记的二抗(1∶1 000)避光孵育2 h;PBS清洗3次,5 min/次;DAPI避光染色3 min;PBS 清洗3次,10 min/次;甘油封片,荧光显微镜下观察拍照。
1.8 牦牛ATG5多克隆抗体的应用
1.8.1牦牛ATG5蛋白在不同年龄牦牛睾丸组织中的表达 取2,4,6,8岁牦牛睾丸组织样品提取总蛋白,与6×蛋白上样缓冲液3∶1混合,100℃水浴10 min 进行蛋白SDS变性。配置10%的分离胶溶液和5%的浓缩胶溶液,进行SDS-PAGE分离蛋白。220 mA冰浴电转1 h,将目标蛋白转移至PVDF膜,使用PBST配制的5%脱脂奶粉溶液封闭2 h;以制备的牦牛ATG5多克隆抗体为一抗(1∶1 000),4℃孵育10 h;TBST 溶液清洗3次,每次10 min;生物素标记的羊抗兔抗体为二抗(1∶7 000),37℃ 孵育1 h;TBST溶液清洗5次,每次5 min,发光显色后进行拍照。通过灰度值分析,以β-actin蛋白作为参照,采用相对定量方法比较不同组ATG5蛋白表达水平。
1.8.2免疫组织化学法检测ATG5蛋白在不同年龄牦牛睾丸组织中的分布 制作常规组织切片,使用0.01 mol/L柠檬酸盐缓冲液进行抗原修复,室温自然冷却后,PBS洗涤3 min×3次;滴加3% H2O2,37℃ 孵育15 min,以阻断内源性过氧化物酶活性,滴加封闭液(A液)室温孵育15 min。滴加制备的牦牛ATG5多克隆抗体(1∶500),4℃ 湿盒孵育过夜,对照组滴加0.02 mol/L PBS替代一抗。滴加二抗(B液),37℃湿盒孵育15 min。滴加C液,37℃湿盒孵育15 min,加DAB试剂显色,苏木精复染,酒精脱水,二甲苯透明,树脂封片,显微镜下观察拍照。

2 结果
2.1 重组质粒pET-32a-ATG5的构建及验证构建的pET-32a-ATG5重组质粒,经限制酶XhoⅠ和ApaBⅠ双酶切形成2条带,分别为1 862 bp(包含ATG5基因片段)和4 853 bp(图1),试验结果符合预期。成功构建的pET-32a-ATG5重组质粒可用于ATG5重组蛋白表达。

M.DL5000 DNA Marker;1.双酶切后的产物片段;2.pET-32a-ATG5重组质粒
2.2 牦牛ATG5重组蛋白原核表达挑选转化平板的6株BL21(DE3)-pET-32a-ATG5单克隆菌,经IPTG 诱导后SDS-PAGE检测,6株单克隆菌均可表达蛋白相对分子质量约为53 kDa的重组蛋白(图2A);经灰度值分析,菌株5的ATG5重组蛋白表达量最高(图2B),可用于牦牛ATG5重组蛋白的纯化。

A.SDS-PAGE检测原核表达的重组蛋白(M.蛋白Marker;1~6.菌株);B.ATG5重组蛋白相对表达水平(1~6.菌株)
2.3 牦牛ATG5重组蛋白的纯化IPTG诱导牦牛ATG5重组蛋白大量表达,并对其进行纯化,经SDS-PAGE鉴定,牦牛ATG5重组蛋白相对分子质量约为53 kDa,(图3A)。透析后,经SDS-PAGE鉴定,目的条带单一无杂带(图3B)。ATG5重组蛋白纯化效果良好,可用于动物免疫。

A.SDS-PAGE检测纯化蛋白(M.蛋白Marker;1.目的蛋白洗脱1;2.目的蛋白洗脱2;3.目的蛋白富集样);B.SDS-PAGE检测纯化透析的蛋白(M.蛋白Marker;1.BSA;2.目的蛋白透析至PBS样品)
2.4 牦牛ATG5多克隆抗体制备
2.4.1Western blot检测抗血清特异性 免疫2只日本大耳白兔后,采集血清。通过Western blot检测,抗血清与牦牛ATG5-His-Trx重组蛋白能发生特异性反应,蛋白相对分子质量为53 kDa(图4),与预期结果一致。对抗血清纯化后得到牦牛ATG5多克隆抗体。

1.重组蛋白1;2.重组蛋白2
2.5 牦牛ATG5多克隆抗体的效价及特异性检测
2.5.1牦牛ATG5多克隆抗体效价检测 通过间接ELISA验证制备的牦牛ATG5多克隆抗体效价,根据D450 nm阳性血清/D450 nm阴性血清>2.1的最大稀释倍数为多克隆抗体的效价,结果显示,所制备的牦牛ATG5多克隆抗体效价大于1∶1 280 000。如表 1、图5所示。

表1 不同稀释度的D450 nm
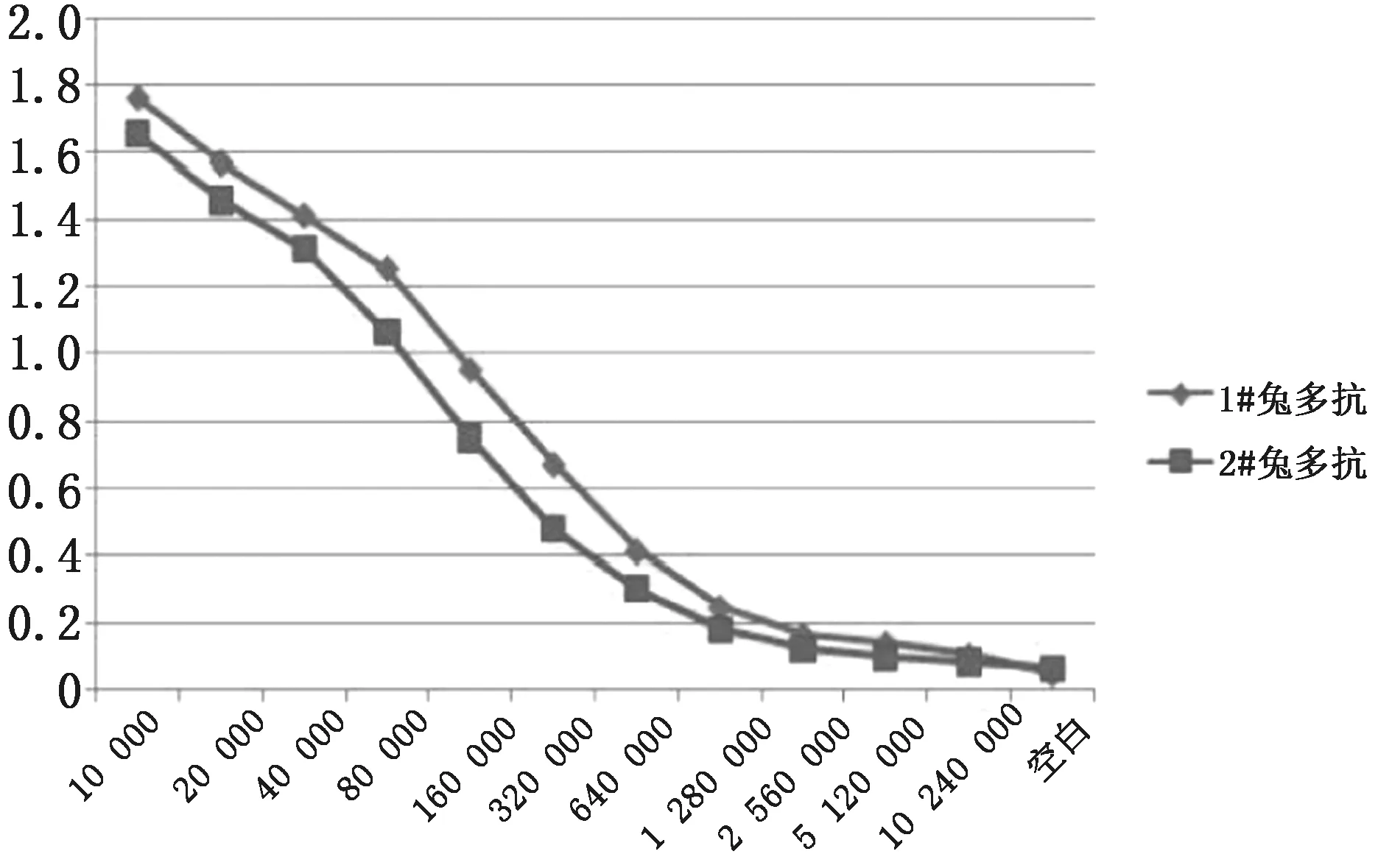
图5 牦牛ATG5多克隆抗体不同稀释度的D450 nm

A.DAPI细胞核染色(蓝色);B.ATG5多克隆抗体特异性染色(红色);C.α-Tubulin内参多克隆抗体特异性染色(绿色);D.合并图像后显色(橙黄色)

A.ATG5蛋白表达检测;B.ATG5蛋白相对表达水平(不同字母表示组间差异显著,P<0.05)
2.5.2免疫荧光检测牦牛ATG5多克隆抗体特异性 通过免疫荧光检测牦牛ATG5多克隆抗体的特异性,结果如图6所示,制备的牦牛ATG5多克隆抗体能与牦牛输卵管上皮细胞中的ATG5蛋白发生特异性结合,ATG5蛋白主要分布在牦牛输卵管上皮细胞细胞核周边及胞质中。
2.6 牦牛ATG5多克隆抗体的应用
2.6.1牦牛ATG5蛋白在不同年龄牦牛睾丸组织中的表达 通过Western blot检测不同年龄组牦牛睾丸组织中ATG5蛋白的表达。ATG5蛋白在各年龄组牦牛睾丸组织中均有表达(图7A),且各组差异显著(P<0.05)。2岁组ATG5蛋白表达水平最低,随着年龄增长ATG5蛋白表达水平随之升高,6岁组ATG5蛋白表达最高。随着牦牛年龄继续增长,8岁组ATG5蛋白表达水平出现下降(图7B)。
2.6.2ATG5蛋白在不同年龄牦牛睾丸组织中的定位 通过免疫组化检测ATG5蛋白在2,4,6,8岁牦牛睾丸组织中的表达,结果如图8所示,2,4,6岁牦牛睾丸组织中,ATG5蛋白表达于支持细胞、精原细胞、初级精母细胞和精子细胞。8岁牦牛睾丸组织中,ATG5蛋白表达于支持细胞和精子细胞。
3 讨论
本研究通过原核表达的方法获得牦牛ATG5重组蛋白,经过蛋白纯化,免疫日本大耳白兔成功获得兔抗牦牛ATG5多克隆抗体。制得的牦牛ATG5多克隆抗体效价大于1∶1 280 000,张加姿等[18]制备的小麦ATG5多克隆抗体效价为1∶25 600,表明本研究所制备的兔抗牦牛ATG5多克隆抗体具有较高的灵敏度。
在原核表达、重组蛋白纯化和抗血清特异性检测过程中,SDS-PAGE测得重组蛋白的相对分子质量约为53 kDa,Western blot测得抗血清与重组蛋白结合的相对分子质量同为53 kDa。在应用多克隆抗体对不同年龄牦牛睾丸组织中ATG5蛋白表达检测过程中,Western blot检测结果显示牦牛ATG5蛋白的相对分子质量约为33 kDa,这与张现伟等[19]对抗人ATG5单克隆抗体的相关研究结果一致。这是由于pET-32a载体自带Trx标签,位于pET-32a载体中目的基因的N-端。ATG5重组蛋白中,Trx蛋白标签的相对分子质量大约为20 kDa。将重组蛋白用于免疫白兔,动物会对ATG5核心蛋白及Trx蛋白标签分别产生相应抗体,但Trx蛋白标签产生的抗体并不与天然蛋白产生特异性结合,并且试验过程中Trx标签也没有对ATG5基因片段的表达造成干扰,不影响ATG5多克隆抗体的特异性。
在验证牦牛ATG5多克隆抗体特异性的免疫荧光检测试验中,结果显示,制备的牦牛ATG5多克隆抗体能够与牦牛输卵管上皮细胞表达的ATG5蛋白发生特异性结合,这与王靖雷等[20]对输卵管组织ATG5蛋白表达的相关研究结果一致。同时,试验还进一步发现ATG5蛋白在牦牛输卵管上皮细胞的表达主要集中在细胞核周边及细胞质中。研究表明,ATG5蛋白能在自噬早期阶段标记正在扩展和生长的吞噬泡[21]。由ATG5蛋白标记的吞噬泡位于内质网(endoplasmic reticulum,ER)的外表面,在快速招募ATG8蛋白之后,最初的吞噬泡开始延伸,其边缘存在着ATG5复合体。吞噬泡的膜一旦闭合,ATG5蛋白则离开该结构,同时,新产生的自噬小泡离开ER[22]。这与本研究在牦牛输卵管上皮细胞免疫荧光检测中得到的结果一致。
本研究通过Western blot和免疫组织化学法以不同年龄牦牛睾丸组织作为研究对象检测ATG5蛋白的表达。Western blot结果显示,ATG5蛋白在2,4,6,8岁牦牛睾丸组织中均有表达。免疫组化结果显示,ATG5蛋白在各年龄段牦牛睾丸支持细胞中表达较为稳定。支持细胞是生精小管中唯一的体细胞,支持细胞不仅通过控制自身数量和功能来调节精子发生,还通过旁分泌作用来滋养支持细胞周围的生殖细胞[23]。同时支持细胞还能通过吞噬作用迅速清除凋亡细胞,维持生精小管环境[24]。这可能表明ATG5蛋白参与精子生成的调节过程。根据灰度值分析不同年龄组ATG5蛋白相对表达水平,2岁组ATG5蛋白表达水平最低,随着年龄增长ATG5蛋白表达水平随之升高,6岁组ATG5蛋白水平显著高于其他年龄组。随着牦牛年龄继续增长,8岁组ATG5蛋白表达水平出现下降。结合免疫组化结果2,4,6岁牦牛睾丸组织中ATG5蛋白分布于支持细胞、精原细胞、初级精母细胞和精子细胞。8岁牦牛睾丸组织中,仅观测到ATG5蛋白位于支持细胞和精子细胞,精原细胞和初级精母细胞未见ATG5蛋白分布。公牦牛的初情期一般为2岁,种公牦牛配种能力最旺盛的时期为3~7岁,7岁以后生殖能力下降。这与本研究结果一致,推测牦牛睾丸组织中ATG5蛋白的表达与公牦牛生殖能力正相关,并且ATG5蛋白在牦牛睾丸组织中的表达极有可能受到下丘脑-垂体-性腺轴及相关激素的调控。
