不同大熊猫扩散地景观格局和植被覆盖变化的比较研究
2023-08-01姜波宇田关胜王翰霖李明明许路冉江洪
姜波宇 , 田关胜, 王翰霖 , 李明明 , 许路 , 冉江洪
(1. 四川大学生命科学学院,生物资源与生态环境教育部重点实验室,成都 610065; 2. 四川大学生命科学学院,四川省濒危野生动物保护生物学重点实验室,成都 610065; 3. 四川申果庄自然保护区管理处,四川 越西 616650)
扩散是动物的基本生活史特征(Walters,2000)。动物通过扩散来达到种群内和种群间个体交换的目的,对防止近亲繁殖、提高种群环境适应能力、扩大种群的分布区具有重要作用(赵洪峰等,2013;陈俪心等,2019)。目前,扩散已成为生态学家关注的重点问题之一(Walters,2000;Clob⁃ertet al.,2009)。
动物扩散到新的生境,面临着继续迁移或定居的选择。生境选择理论通常认为,个体通过做出最优的定居决策,在异质性景观中选择最高质量的栖息地,最大化其适合度(Fretwell,1969)。而影响栖息地质量的因子很多,景观格局和植被覆盖均是影响栖息地质量的重要因子(徐文茜,2016)。景观格局是从宏观尺度反映栖息地质量的主要方法指标(李向阳等,2009;刘艳萍等,2012),很大程度上影响着动物种群的分布(范隆庆等,2010)。此外,植被覆盖也会影响栖息地质量,一般植被覆盖越好,区域生态环境质量也越好(王琦等,2019)。同时,植被覆盖及其动态变化在一定程度上会潜在影响动物栖息地的分布与变化(卢学理等,2008;赵海迪等,2014)。
大熊猫Ailuropoda melanoleuca是世界生物多样性保护的旗舰物种。目前孤立地分布在我国四川、甘肃、陕西的6 大山系中,其中,凉山山系是大熊猫分布的最南端(和梅香等,2018)。有关大熊猫微生境和宏生境选择的研究较多(Qiet al.,2009,2011;和梅香等,2018;李杨倩等,2018),但对大熊猫扩散的研究却鲜有报道,目前仅有通过群体遗传学数据分析得出大熊猫是偏雌性扩散(Zhanet al.,2007;Huet al.,2010),而对扩散大熊猫的宏生境选择还未知。
四川省第四次大熊猫调查结果显示,凉山山系越西县东北部的梅花乡-白果乡区域和雷波县西部的拉咪乡-长河乡区域是2012 年调查发现的大熊猫新分布区,其在2001 年第三次大熊猫调查时均无大熊猫分布(四川省林业厅,2015)。通过对这2 个区域的调查和长期监测,发现截至2021 年越西县梅花乡-白果乡区域一直有大熊猫活动痕迹,而雷波县拉咪乡-长河乡区域在2014—2021 年未发现大熊猫痕迹,这为比较不同大熊猫扩散地景观格局和植被覆盖变化,以及大熊猫扩散的宏生境选择提供了机会。
本文利用3S技术与景观生态学的数量分析方法定量对比分析了2 个扩散区域的景观格局和植被覆盖在近10 年间的时空变化特征,探究景观格局和植被覆盖变化对大熊猫宏生境选择的潜在影响,以期了解可能影响扩散大熊猫宏生境利用的主要因子,识别可能影响大熊猫扩散利用的关键环境参数,为大熊猫栖息地的保护与恢复提供科学依据。
1 研究区域概况
研究区域位于四川省凉山山系,根据2012 年全国第四次大熊猫调查对大熊猫栖息地的范围划分,以山脊或道路为界(李云飞等,2015),越西区域:越西县东北部的梅花乡-白果乡(102.61°~102.70°E,28.73°~28.87°N),海拔1 517~3 579 m,面积80.22 km2,第四次大熊猫调查发现大熊猫痕迹点18 个,2021 年和2022 年仍然发现有大熊猫活动痕迹;雷波区域:雷波县南部的拉咪乡-长河乡(103.27°~103.42°E,28.17°~28.44°N),海拔1 437~3 471 m,面积173.74 km2,第四次大熊猫调查发现大熊猫痕迹点5个,2014—2021年未发现大熊猫活动痕迹(图1)。
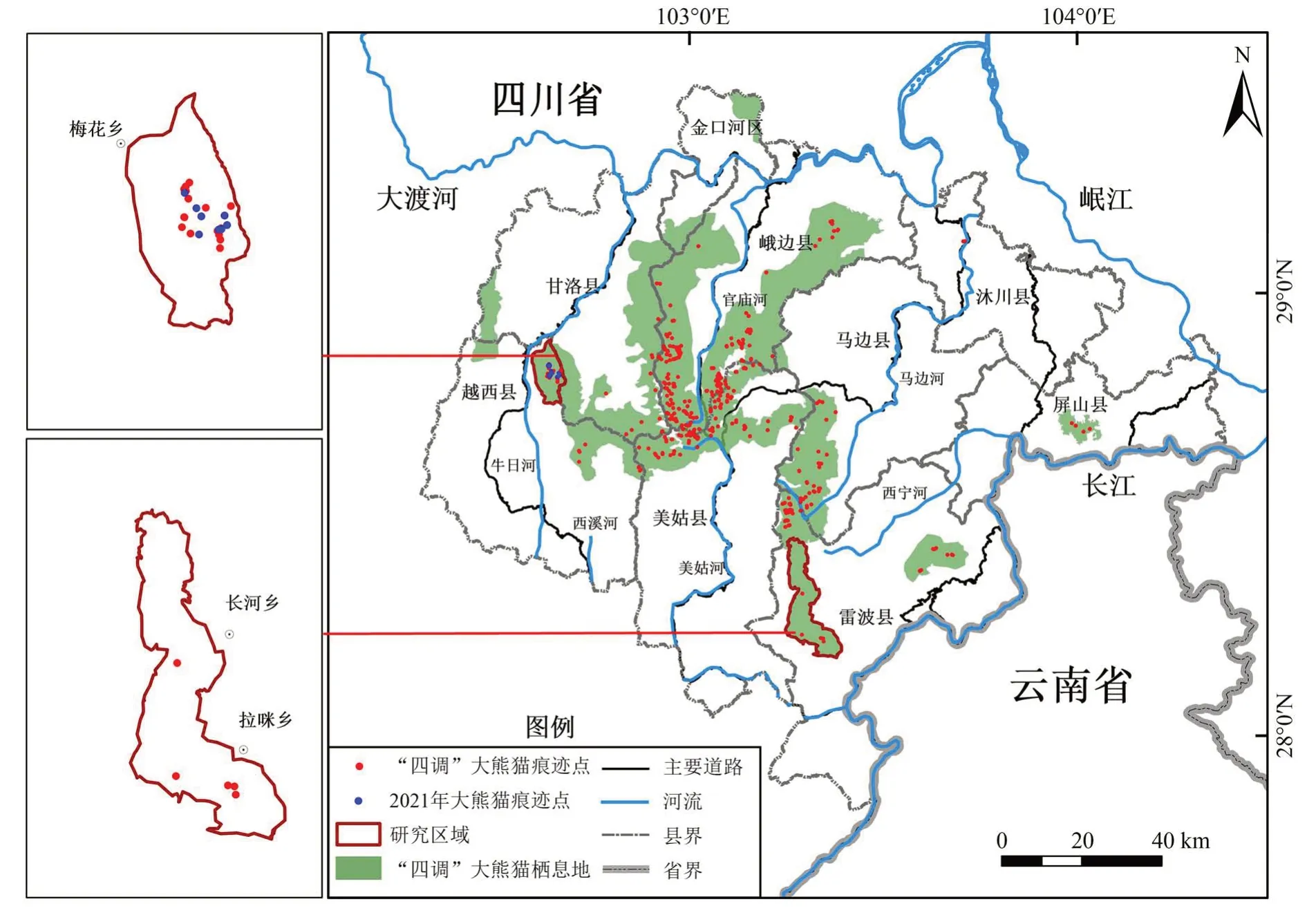
图1 研究区域Fig. 1 Study area
2 研究方法
2.1 野外调查方法
2.1.1 植被调查 调查样线按照全国第四次大熊猫调查标准布设,在样线上合适位置设置植被调查样方,大小为20 m×20 m,记录植被类型和地理坐标(饶佳等,2018),用于遥感影像的解译及之后的精度评价。2021 年6 月在越西区域和雷波区域分别设置了22 条和34 条样线,70 个和105 个植被样方。
2.1.2 动物调查 在进行植被调查的同时进行大熊猫痕迹调查,即发现大熊猫痕迹点时,记录痕迹点类型及其经纬度、海拔等信息。在2021年6月、8月、12月和2022年4月进行了4次调查。
2.2 遥感影像数据获取
遥感影像数据选择Landsat8卫星影像,数据来自地理空间数据云(http://www.gscloud.cn)和美国地质勘探局USGS(https://earthexplorer.usgs.gov)。选取研究区域2011 年和2021 年植被生长季且云量低于5%的TM 影像。数据波段空间分辨率均为30 m。
在ENVI5.3 中对2 个研究区域2011年和2021年的Landsat8 遥感影像进行解译。首先对影片进行预处理,具体过程包括大气校正、几何精校正、辐射定标、拼接与裁剪、监督分类与非监督分类相结合等。监督分类主要使用最大似然分类方法,最终将研究区域内土地覆盖类型划分为针叶林、阔叶林、灌丛、草甸、农业用地和建筑用地6类。
选择Cohen 提出的Kappa 系数法(Cantor,1996),并结合实地植被样方数据和奥维卫星影像图,由于研究区域面积较小,每一类别分别选取尽可能多的点用于精度评价,以减少由于影像分辨率差异和几何纠正引起的误差,若Kappa系数大于0.8,总体精度均达到85%以上,表明解译精准度较高,解译结果与真实情况有较强的一致性(Manandharet al.,2009)。2011 年解译结果通过全国第四次大熊猫调查数据库中记录的植被样方数据进行精度评价,2021 年解译数据通过2021 年野外调查记录的样方数据进行精度评价。
2.3 景观指数分析
用Fragstats 4.2.1 分别计算研究区域的景观格局指数,包括斑块面积、平均斑块面积、相似邻近比度、聚集度指数、景观形状指数、香农多样性指数和破碎化指数。采用Mann-WhitneyU检验分别对2 个区域各景观类型2011年和2021年的斑块面积进行差异分析,统计分析在SPSS 25中进行。
2.4 植被覆盖分析
归一化植被指数(normalized differential veg⁃etation index,NDVI)是反映区域植被动态变化最敏感的指标之一(王小霞等,2020),对植被覆盖度检测幅度较宽,具有良好的相位和空间适应性,被广泛应用于反映区域尺度的植被覆盖度状况(盖永芹等,2009)。
为降低其他干扰因素的影响,选择植被生长物候期(4—10 月)的数据,以2011 年和2021 年生长期内森林的NDVI 均值代表当年的森林覆盖度。NDVI 数据来源于美国国家航空航天局NASA(https://ladsweb. modaps. eosdis. nasa. gov/search/)MODIS 数据产品的MOD13Q1 数据集,空间分辨率为250 m×250 m,时间分辨率为16 d。运用MODIS Reprojection Tools 对原始数据进行格式和投影转换,然后在ArcGIS 10.2 用最大值合成法合成生长期年NDVI数据(Holben,1986)。
根据四川省第四次大熊猫栖息地评价图层,将适宜栖息地作为高质量栖息地(国家林业和草原局,2021),分别计算2011 年和2021 年研究区域和高质量栖息地的NDVI,栅格大小为250 m×250 m。NDVI年际变化率(K)通过2011年和2021年各像元的NDVI 值计算得到:K=(NDVI2021-NDVI2011)/10。其中,K<−0.005 表示森林覆盖度呈减少趋势,−0.005≤K≤0.005 表示森林覆盖度基本不变,K>0.005 表示森林覆盖度呈增加趋势(马晓勇等,2019)。采用Mann-WhitneyU检验分别对2个区域及其高质量栖息地各栅格10 年间的NDVI 值进行显著性差异分析,统计分析在SPSS 25中进行。
3 研究结果
3.1 分类及精度评价结果
分类结果显示,越西区域2011 年和2021 年的解译总体精度分别为89.26%和87.59%,Kappa 系数分别为0.823和0.839;雷波区域2011年和2021年的解译总体精度分别为86.23%和87.77%,Kappa系数分别为0.817和0.847(图2)。

图2 2011年和2021年越西区域、雷波区域的景观格局Fig. 2 Landscape patterns of Yuexi area and Leibo area in 2011 and 2021
3.2 景观斑块类型尺度的变化
3.2.1 景观斑块类型面积变化 研究区域的主要景观类型均为森林,包括针叶林和阔叶林,是整个景观格局的基质;灌丛和草甸面积均不大;农业用地和建筑用地占比很少。10 年间,越西区域针叶林面积减少7.18%,阔叶林面积增加2.97%,森林面积总体减少;雷波区域针叶林面积增加5.72%,阔叶林面积增加5.96%,森林面积总体增加(表1)。
3.2.2 景观斑块类型破碎化变化 2011—2021年,越西区域针叶林和阔叶林的平均斑块面积、相似邻近比度、聚集度指数均增加,且针叶林的平均斑块面积增加明显;森林整体破碎化情况减弱。雷波区域针叶林的平均斑块面积增加,但相似邻近比度和聚集度指数均减小;阔叶林的平均斑块面积减小,相似邻近比度和聚集度指数也有一定程度减小;森林整体破碎化情况增强。
Mann-WhitneyU检验结果表明,2011—2021年,越西区域除草甸和建设用地的平均斑块面积无显著变化(P>0.05)外,针叶林极显著增加(P<0.01),其他景观斑块类型显著增加(P<0.05);雷波区域的阔叶林、灌丛和草甸的平均斑块面积极显著减少(P<0.01),针叶林和农业用地显著增加(P<0.05),建筑用地显著减少(P<0.05)(表2)。

表2 2011年和2021年研究区域不同景观类型的平均斑块面积、相似邻近比度和聚集度指数Table 2 Mean patch size, percentage of like adjacencies and aggregation index of different landscape types in the two study areas between 2011 and 2021
3.3 景观尺度的变化
2011—2021 年,越西区域的景观破碎化指数和景观形状指数均减小,香农多样性指数增加;雷波区域的景观破碎化指数和景观形状指数均增加,香农多样性指数降低(表3)。

表3 2011年和2021年研究区域不同景观类型的景观形状指数、香农多样性指数和破碎化指数Table 3 Landscape shape index, Shannon’s diversity index and fragmentation index of different landscape types in the two study areas between 2011 and 2021
3.4 区域整体和高质量栖息地的植被覆盖度对比
2011—2021 年,2 个区域绝大部分栅格的NDVI 值>0.6,部分栅格的NDVI 值>0.8。10 年间,79.85%越西区域的NDVI值基本保持不变,17.33%增加,高质量栖息地的NDVI 值平均增加0.025;91.31%雷波区域的NDVI 值基本保持不变,7.53%增加,高质量栖息地的NDVI 值平均增加0.019。2011—2021 年,2 个区域高质量栖息地的NDVI 值均显著增加,但NDVI 值的增加量没有显著差异(图3)。

图3 2011—2021年越西区域和雷波区域植被覆盖(NDVI值)的变化Fig. 3 Vegetation coverage (NDVI) changes in Yuexi area and Leibo area from 2011 to 2021
4 讨论
本研究结果显示,2 个区域森林面积的变化幅度都相对较小。2011—2021 年,越西区域针叶林面积有一定减少,主要是由于针叶树种被当地社区居民作为木材和薪材采伐所致;其他景观类型的转换主要是由放牧、耕地开垦、新农村基础设施建设等活动所致。雷波区域的变化主要源自植树造林导致的灌丛和草地面积减少,森林面积增加,新农村和基础设施建设导致了建设用地增加。
2011—2021 年,雷波区域整体森林破碎化程度增加,主要是由于人工新造林及植被自然恢复更替等导致针叶林和阔叶林斑块镶嵌较多;而2001 年前森林采伐和农垦活动频繁,致使林相退化,以次生林为主(戴波等,2014)。而次生林是由多个先锋树种形成不同的斑块,导致区域异质性增加,形成的斑块面积也较小,森林群落的演替会使区域破碎化程度加剧(张会儒等,2009)。越西区域则主要是部分针叶林转化为阔叶林,导致区域内针叶林、阔叶林斑块分布更规则、聚集,整体森林破碎化程度降低。
2011—2021 年,越西区域景观受人类活动干扰程度减少,景观整体破碎化程度减弱;而雷波区域受人类活动干扰程度增加,景观整体破碎化程度增加。对于以森林为主要景观类型的区域,植被的次生演替可以使景观结构趋于复杂,景观丰富度提高,多样性指数提高;而诸如采伐、开荒等人类活动往往使景观类型变得单一、丰富度下降、多样性指数降低(张爽等,2004)。越西区域植被次生演替情况更好,植被恢复情况更好,而雷波区域植被次生演替情况较差,且人类活动更频繁,这与2 个区域景观形状指数得出的结论一致。产生这种结果的原因可能是,雷波区域村庄较多且分散,居民的农牧活动对整体景观影响较大;且过去10 年间人工林种植较多,种植类型较为单一,也使其植被次生演替情况较差,多样性指数较低。
2 个区域在2011 年和2021 年均为植被覆盖良好区域(NDVI>0.6)(王晓利等,2013)。2011—2021年,2 个区域中只有部分区域的NDVI 增加,而大部分区域维持不变。越西区域的增加部分大多属于天然林,减少区域主要位于居民区附近,由开垦农田、放牧、林木砍伐等所致。雷波县的增加区域主要位于人工造林区域,减少区域主要是居民点周边及草地区域,由林木砍伐及放牧等活动所致。
栖息地破碎化和植被覆盖变化是宏观尺度上对动物栖息地选择的两大重要影响因素(杨少文,2015)。对于大熊猫这样的大型哺乳动物,栖息地破碎化和植被覆盖变化影响其分布的现象尤为明显(白文科等,2017;朱若柠等,2019)。森林和整体景观破碎化使大熊猫的活动范围减小,生存环境孤立,影响大熊猫的扩散行为(李国春,2017;陈文德等,2021)。本研究发现,扩散大熊猫对宏生境的选择也与森林和景观破碎化有关,偏向于选择森林(尤其是针叶林)和整体景观破碎化较低的生境,这与张爽等(2004)的研究结果类似。而植被覆盖度与大熊猫的隐蔽选择、栖息地选择紧密联系,大熊猫对隐蔽条件的要求可理解为对植被(尤其是乔木)覆盖度的选择及对各种干扰的趋避(孙亮等,2022),其选择具有较高乔木郁闭度的发育成熟的浓密林冠下的栖息地(Wanget al.,2022),较大的乔木郁闭度可为其提供良好的隐蔽条件(孙飞翔等,2013)。但同时,林下竹子的生长也需要一定的光照条件,较低的郁闭度和乔木密度(植被覆盖度)可以满足林下竹子生长对光照的需求(李爽等,2017)。本研究并没有发现植被覆盖的提升对大熊猫的宏生境选择产生积极影响,可能是因为在2 个区域植被覆盖度自身基数较高的情况下,植被覆盖度的增加并不会使生境的适合度产生明显的提高。
在宏观尺度上,大熊猫扩散后居留的越西区域相较其短期停留后又离开的雷波区域,森林和整体景观破碎化程度均相对较低,景观受人为活动影响较小,大熊猫主要活动的针叶林平均斑块面积较大,同时区域的香农多样性指数也较高。这可能预示着大熊猫对新扩散区域的森林和整体景观破碎化、针叶林的平均斑块面积具有一定的选择性,同时可能偏好森林管护好、人为活动影响小、植被快速演替恢复的区域。这提示在大熊猫栖息地保护与恢复时,需要注重栖息地景观结构的恢复,减少森林景观破碎化程度,扩大其主要活动栖息地的斑块面积,同时加强管控措施减少干扰,促进植被的健康恢复。
影响大熊猫宏生境选择的主要因素包括地理环境、森林群落结构、主食竹和干扰等(何敏等,2018),本研究仅涉及到影响大熊猫扩散的地理环境因素和森林群落结构因素中的一部分。影响大熊猫扩散居留的还有其生物学特征,如繁殖等生理因素(魏辅文,2018),并不完全由外界条件决定。因此,后续研究还需要结合更多类型数据,从多个方面进行综合分析,以进一步了解扩散大熊猫对宏生境的选择,科学系统地指导大熊猫的保护与栖息地恢复工作。本结果可为认识扩散大熊猫的宏生境选择提供有益的参考,对大熊猫栖息地恢复具有积极作用。
致谢:感谢四川省林业和草原局提供的四川省第四次大熊猫调查资料;感谢越西县林业和草原局、雷波县林业和草原局和越西申果庄自然保护区、雷波嘛咪泽自然保护区等单位与相关人员给予的帮助。感谢李雨航师兄、刘雨欣师姐、夏林超等人对数据收集与分析、制图、论文修改的帮助。
