籽瓜多糖对益生菌生长促进效应及其结构表征
2022-10-18常雪花钱雅雯王振菊魏佳吴斌张政
常雪花,钱雅雯,王振菊,魏佳,吴斌,张政*
(1.巴音郭楞职业技术学院生物工程学院 新疆 库尔勒 841000;2.新疆农业科学院农产品贮藏加工研究所,新疆 乌鲁木齐 830091;3.新疆农产品加工与保鲜重点实验室,新疆 乌鲁木齐 830091)
籽瓜(Citrullus lanatus ssp.Vulgaris var.megalaspermus Lin et Chao),属葫芦科(Cucurbitaceae)西瓜属(Citrullus genus),是新疆重要的特色农作物之一。随着新疆优势农产品区域化布局的进一步推进,籽瓜产业迅速发展。2020年,籽瓜播种面积达84 990万m2,产量为195 451 t,其产量仅次于番茄和辣椒[1]。长期以来,取籽加工为籽瓜最主要产品形式,超过籽瓜总质量90%以上的瓜瓤和瓜皮被直接废弃,其副产物综合利用率较低,这导致籽瓜产业整体产品附加值不高,严重阻碍了籽瓜产业升级转型和可持续发展[2-3]。
近年来,多糖在功能性食品领域受到越来越多的关注。以益生元为导向的多糖筛选和开发已逐渐成熟,成为调节肠道菌群以维持身体健康的主要策略之一[4-5]。枸杞多糖具有益生元特性,能增强肠道微生物群,促进嗜酸乳杆菌和长双歧杆菌的体外生长[6]。此外,Ouyang等[7]从山药中分离出的 YPN、YPN-Ⅰ、YPN-Ⅱ和YPN-Ⅲ4个多糖组分能明显促进嗜热链球菌的生长。课题组前期利用响应面法优化籽瓜多糖的超声辅助水提取工艺,提取出的籽瓜多糖含量达到6.03%[8]。这表明籽瓜可作为一种多糖的生物来源。籽瓜粗多糖能够缓解细胞氧化应激损伤[9]。从“内蒙黑中片”籽瓜中分离纯化的多糖SI具有较好的抗氧化能力,能抑制酵母菌的体外生长[10]。目前,人们对籽瓜多糖功能性作用的认知主要局限于其抗氧化活性和抑菌活性,这不能充分和科学地评价籽瓜多糖的功能特性。籽瓜多糖是否能与枸杞和山药多糖等一样,作为一种潜在的益生元仍需进一步研究。
本文以新疆籽瓜为试验材料,对籽瓜多糖进行分离、纯化,比较各组分对益生菌生长影响的差异性,明确最优组分的一级结构,为籽瓜多糖生物活性与结构表征间构效关系以及籽瓜高附加产品的开发提供理论基础。
1 材料与方法
1.1 材料与试剂
籽瓜于2021年8月采摘于新疆焉耆县籽瓜种植园。
青春双歧杆菌、长双歧杆菌、婴儿双歧杆菌、嗜酸乳杆菌:中国普通微生物菌种保藏管理中心;DEAE纤维素DE-52填料:上海远业生物科技有限公司;石油醚、氢氧化钠、硫酸:天津市北联精细化学品开发有限公司;溴化钾、氯化钠、磷酸二氢钠、磷酸氢二钠、硫酸铁、葡萄糖:福晨(天津)化学试剂有限公司;牛肉浸粉、大豆蛋白胨、胰蛋白胨、酵母浸粉、肝浸粉:青岛海博生物技术有限公司。以上试剂均为分析纯。
1.2 仪器与设备
1260高效液相色谱仪:安捷伦科技(中国)有限公司;GCMS-QP2010 SE气相色谱-质谱联用仪:岛津(上海)实验器材有限公司;FTIR-650傅里叶变换红外光谱仪:天津港东科技股份有限公司;Bruker AV-600MHz核磁共振仪:布鲁克(北京)科技有限公司;GJ-25C冷冻干燥机:北京四环科学仪器厂有限公司;RE100-Pro旋转蒸发仪:美国SCILOGEX公司;HH-2恒温水浴锅:江苏金坛市荣华仪器制造有限公司;UV-2600紫外可见分光光度计、PHS-3C pH计:上海耶茂仪器设备有限公司;LDZX-30KB高压灭菌锅:上海鼎谦生物科技有限公司;圆底立式厌氧培养袋:青岛海博生物技术有限公司;DHG-9023A鼓风干燥箱、SHP-250生化培养箱:上海精宏实验设备有限公司。
1.3 试验方法
1.3.1 籽瓜多糖的提取和分离
将籽瓜去翠衣皮、切块、去籽、留瓜瓤,经40℃低温干燥48 h后粉碎,经100目筛网过滤得到籽瓜干粉。将籽瓜干粉按原料:石油醚为1∶3(g/mL)的料液比加入石油醚原液,室温(20±5)℃下浸泡12 h后,采用旋转蒸发仪浓缩体积,回收石油醚。重复2次,除去脂溶性杂质,最后放入40℃烘箱内干燥,得到籽瓜干粉原料。
采用课题组前期优化的超声辅助水提法提取籽瓜多糖[6]。籽瓜多糖分离参考Zhu等[11]的方法,略有修改。取1g籽瓜干粉溶解在30mL蒸馏水中,42℃、220W超声辅助提取2 h后,12 000 r/min离心10 min。将上清液缓慢倒入DEAE纤维素色谱柱中。随后调整流速至15 mL/min,分别用4组溶剂(3倍柱体积的蒸馏水、0.2 mol/L NaCl、0.5 mol/L NaCl和 2.0 mol/L NaCl)洗脱。采用苯酚硫酸法在490 nm处检测洗脱液吸光度。
1.3.2 菌株培养
称取牛肉浸粉3.0 g,可溶性淀粉0.5 g,L-半胱氨酸0.5 g,大豆蛋白胨、胰蛋白胨各5.0 g,酵母浸粉、肝浸粉各 5.0 g,葡萄糖 10.0 g,NaCl、K2HPO4、KH2PO4各1.0 g以及 FeSO4·7H2O 0.01 g,用蒸馏水溶解,定容至1 000 mL并调节pH值为7.2,再加入20.0 g琼脂,即为双歧杆菌培养基。
取蛋白胨、牛肉浸粉各10.0 g,酵母提取物5.0 g,葡萄糖20.0 g,K2HPO42.0 g和MgSO40.1 g溶解后,定容至1 000 mL并调节pH值为6.5,再加入15.0 g琼脂,即为嗜酸乳杆菌培养基。
配制的液体培养基在121℃高压灭菌30 min后,置于灭菌厌氧培养袋中,形成厌氧环境。使用0.3 mL无菌水对不同菌种冻干粉进行溶解,并在培养基中接种。将接种后的培养基放入厌氧培养袋中迅速加入厌氧产气包,并且快速封口。随后置于37℃培养箱中培养48 h。通过划线平板法,对培养完毕的菌液进行接种,37℃环境下进行厌氧培养48 h。抽取单菌置于灭菌处理后的液体培养基,再于37℃培养48 h。
1.3.3 益生菌的增殖和产酸测定
益生菌的增殖测定参考包晓玮等[12]的方法,略有修改。试管中加入10 mL培养基,在121℃下高温灭菌20 min。向培养基中添加籽瓜多糖组分使其终浓度为2%。对活化状态的试验菌株进行接种,37℃厌氧密封条件下培养48 h,以1.3.2中配制的培养基为对照(CK)。测定各液体培养基的pH值,从而探究籽瓜多糖对试验菌株增殖和产酸影响的差异。
1.3.4 籽瓜多糖对益生菌生长速率的影响
益生菌生长速率的测定参考Ouyang等[7]的方法,稍作修改。在试管内倒入10 mL培养基,再分别加入200 mg的SMP-1、SMP-2和SMP-3,得到3种含2.0%多糖组分培养基。经蒸馏水溶解,4 000 r/min下离心10 min,取上清液。在121℃下对该培养基进行高温灭菌20 min。对活化状态的4种菌株依次进行接种,厌氧条件下培养48 h。以1.3.2中配制的培养基为对照(CK)。分别在 0、4、8、16、24、32、40 和 48 h 测定培养液的吸光度(660 nm)。绘制4株菌在添加籽瓜多糖的培养基的生长曲线,评价4种肠道益生菌在多糖环境下的生长速率。
1.3.5 分子量的测定
分子量的测定参考Zhou等[13]的方法,略有改动。将100 mg粗多糖溶解至3 mL蒸馏水中,12 000 r/min离心10 min,取多糖溶液上清液进行上样,利用多糖凝胶纯化系统结合示差检测器进行纯化样品,并且在线检测收集。将收集液通过旋转蒸发仪进行浓缩、冷冻干燥,得到凝胶柱分离纯化过的多糖用于分子量测定。
取适量SMP-1配制成5 mg/mL溶液,12 000 r/min离心10 min,上清液用0.22 μm微孔滤膜过滤,然后将样品转置于1.8 mL进样瓶中,进样量20 μL。以不同相对分子质量的葡聚糖[分子质量分别为1 152、11 600、23 800、48 600、80 900、148 000、273 000、409 800 重均分子量(Mw)]作为标准品,作标准曲线,测定多糖的相对分子质量,得到峰位分子量、重均分子量、数均分子量校正曲线。根据标准品曲线,计算出样品的分子量大小。
1.3.6 多糖的单糖组成分析
多糖的单糖组成分析参考Xu等[14]的方法,采用高效液相色谱仪进行分析。
1.3.7 红外光谱分析
精密称取SMP-1 2 mg和溴化钾200 mg,压制成片,空白采用溴化钾粉末压片而成。分别置于FTIR-650傅里叶变换红外光谱仪中进行扫描记录。
1.3.8 甲基化分析
甲基化分析参考Xu等[14]的方法。采用GCMS-QP-2010 SE气相色谱质谱联用仪对乙酰化产物进行测定。
气相色谱质谱联用(gas chromatograph-mass spectrometer,GC-MS)条件:色谱柱30 m×0.25 mm×0.25 μm;升温条件为起始120℃,从3℃/min逐渐升温,直到250℃;进样口和检测器温度均为250℃,以氦气为载气,使流速达1 mL/min。
1.3.9 核磁共振分析
称取SMP-1样品50 mg,将其溶于0.5 mL重水中并冷冻干燥,重复3次,以充分交换活泼氢。然后将冷冻干燥后的样品再用0.5 mL重水溶解。采用Bruker AV-600 MHz核磁共振(nuclear magnetic resonance,NMR)采集1H NMR谱、13C NMR谱、DEPT 135谱、氢-氢相关谱(H-H correlation spectroscopy,HH-COSY)、异核单量子关系谱(heteronuclear singular quantum correlation,HSQC)、异核多键相关谱(heteronuclear multiple bond correlation,HMBC)和核欧沃豪斯效应谱(nuclear overhauser effect spectroscopy,NOESY)分析多糖结构。
1.4 数据处理
试验数据采用Microsoft excel 2010和SPSS 22.0进行图形处理和分析。单个试验均设置3个重复,采用均值±标准差形式表示试验结果。
2 结果与分析
2.1 籽瓜多糖的分离
籽瓜多糖的洗脱曲线见图1。
不同多糖组分的荷电性质存在差异,通过提高洗脱溶剂(NaCl)的离子强度从离子交换器中分离多糖组分。DEAE纤维素DE-52是分离多糖的常用介质,适用于各种中性和酸性多糖的分离。由图1可知,籽瓜多糖在使用DEAE纤维素DE-52分离后,观察到3个洗脱峰,并分别命名为SMP-1、SMP-2和SMP-3。收集3个组分,浓缩、脱盐和冻干以进行下一步试验。
2.2 籽瓜多糖组分对益生菌产酸的影响
培养基pH值的变化可以间接反映益生菌的生长状况,青春双歧杆菌、婴儿双歧杆菌、长双歧杆菌和嗜酸乳杆菌培养基pH值变化见图2。
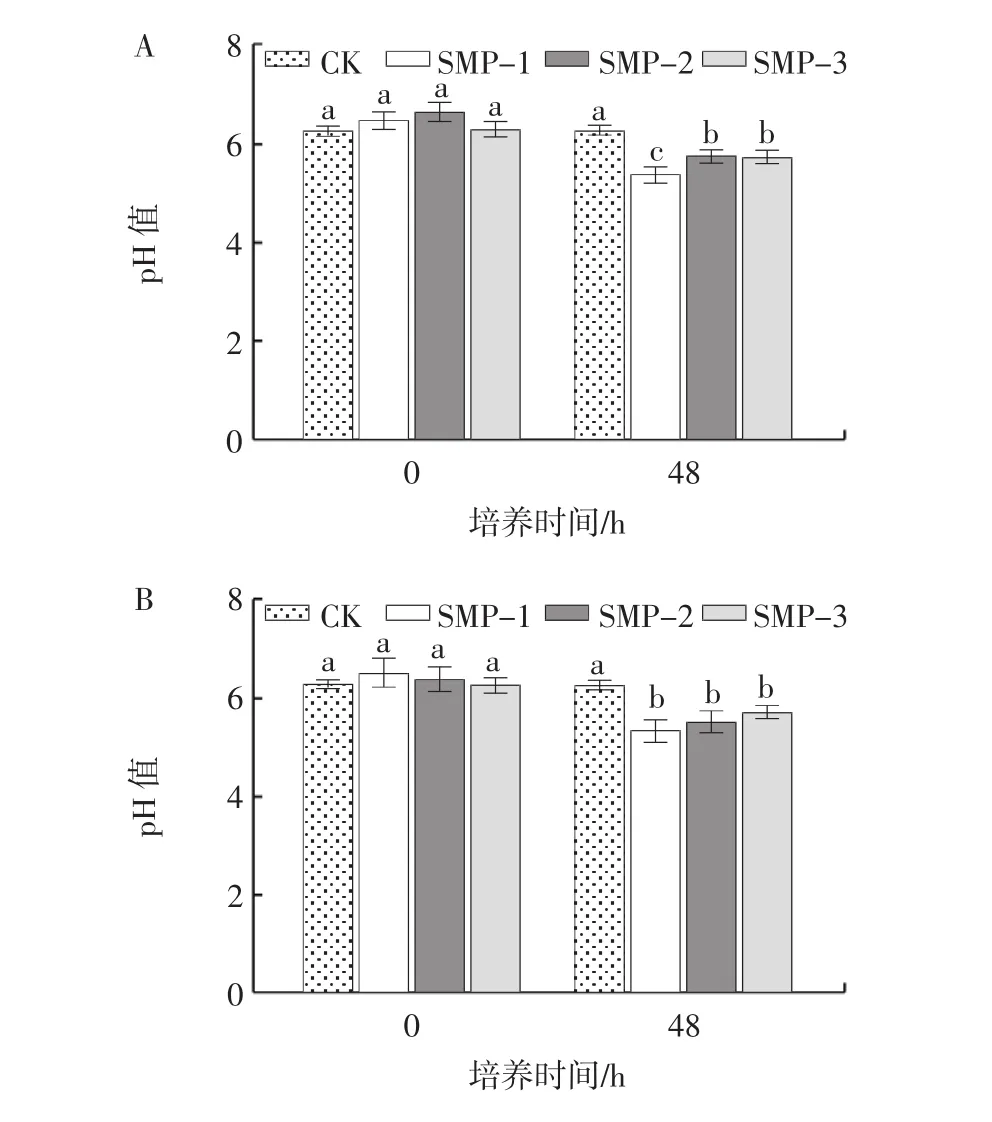
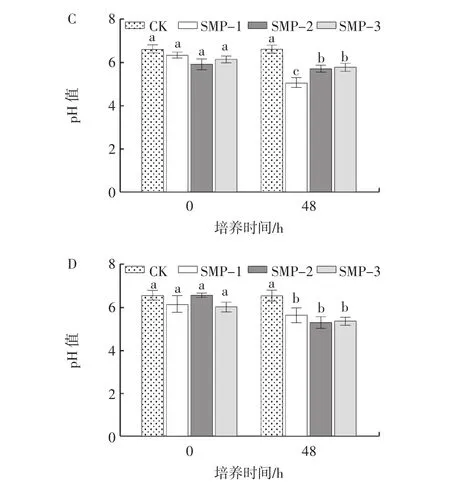
图2 籽瓜多糖组分对青春双歧杆菌、婴儿双歧杆菌、长双歧杆菌和嗜酸乳杆菌培养基pH值的影响Fig.2 Effects of seed melon polysaccharides on media pH of Bifidobacterium adolescentis,Bifidobacterium longum,Bifidobacterium infantis and lactobacillus acidophilus
由图2可知,随着培养时间的延长,SMP-1、SMP-2和SMP-3组的pH值均逐渐下降,且3个籽瓜多糖组分均显著促进了益生菌的增殖。对于青春双歧杆菌和长双歧杆菌,培养48 h时,SMP-1、SMP-2和SMP-3组的pH值均显著低于对照组(p<0.05),且SMP-1的增殖作用优于SMP-2和SMP-3组(p<0.05)。而在接种婴儿双歧杆菌和嗜酸乳杆菌培养48 h时,SMP-1、SMP-2和SMP-3组间pH值无显著差异(p>0.05)。结果表明,籽瓜多糖更好地被益生菌利用,促进益生菌的增殖。综合考虑,选择SMP-1进行后续试验。
2.3 籽瓜多糖对益生菌生长速率产生的影响
600 nm波长下的OD值是反映益生菌生长状况的方法之一,OD值与益生菌生长呈正比。籽瓜多糖组分SMP-1、SMP-2和SMP-3对青春双歧杆菌、婴儿双歧杆菌、长双歧杆菌和嗜酸乳杆菌生长的促进作用如图3所示。

图3 籽瓜多糖组分对青春双歧杆菌、婴儿双歧杆菌、长双歧杆菌和嗜酸乳杆菌生长速率的影响Fig.3 Effects of seed melon polysaccharides on the growth rate of Bifidobacterium adolescentis,Bifidobacterium longum,Bifidobacterium infantis and lactobacillus acidophilus
在添加籽瓜多糖SMP-1、SMP-2和SMP-3的培养基中,青春双歧杆菌培养48 h后,OD600nm维持稳定(图3A),这表明青春双歧杆菌已处于相对平稳的生长状态。8 h后,SMP-1对青春双歧杆菌的促生长作用比各组分差异都显著(p<0.05)。婴儿双歧杆菌培养32 h后SMP-2组OD600nm达到最大值,而嗜酸乳杆菌SMP-3组OD600nm最高(图3B和图3D),且培养结束时,婴儿双歧杆菌SMP-2组的OD600nm和嗜酸乳杆菌SMP-3组均高于对照和SMP-1组(p<0.05)。对于长双歧杆菌,SMP-1组的OD600nm在16 h后始终显著高于 SMP-2和 SMP-3组(p<0.05),说明 SMP-1对长双歧杆菌的促生长作用要优于SMP-2和SMP-3(图3C)。
综上所述,SMP-1、SMP-2和SMP-3对益生菌的促进作用的影响存在差异,但3个组分均能促进4个益生菌的生长繁殖。结果表明,籽瓜多糖可能是一类潜在的益生元。
2.4 相对分子量分析
根据多糖组分对益生菌生长影响的测试结果,选择SMP-1进行进一步的结构表征。SMP-1高效凝胶渗透色谱如图4所示。

图4 籽瓜多糖SMP-1高效凝胶渗透色谱分析Fig.4 High-performance gel permeation chromatography profiles of SMP-1 of seed melon polysaccharides
由图4可知,SMP-1有3个吸收峰,有进一步分离纯化的空间,其分子量为97 403 Da。
2.5 单糖成分分析
SMP-1单糖成分分析见图5。籽瓜多糖SMP-1的单糖组成和物质的量百分比见表1。

图5 单糖标准品和籽瓜多糖SMP-1的单糖组成Fig.5 Monosaccharide composition of standard and SMP-1

表1 籽瓜多糖SMP-1的单糖组成和物质的量百分比Table 1 Monosaccharide composition and molar percentage of SMP-1 of seed melon polysaccharides
由图5可知,单糖标准品的出峰顺序分别为鼠李糖、岩藻糖、阿拉伯糖、木糖、甘露糖、葡萄糖、半乳糖。各单糖标准品分离效果明显,峰形尖锐,可用于后续SMP-1单糖组成比对分析。比对结果如表1所示,在7个单糖中,SMP-1多糖仅含有阿拉伯糖和半乳糖,其物质的量百分比分别为0.271%和0.729%。
2.6 红外图谱分析
籽瓜多糖SMP-1的红外光谱见图6。
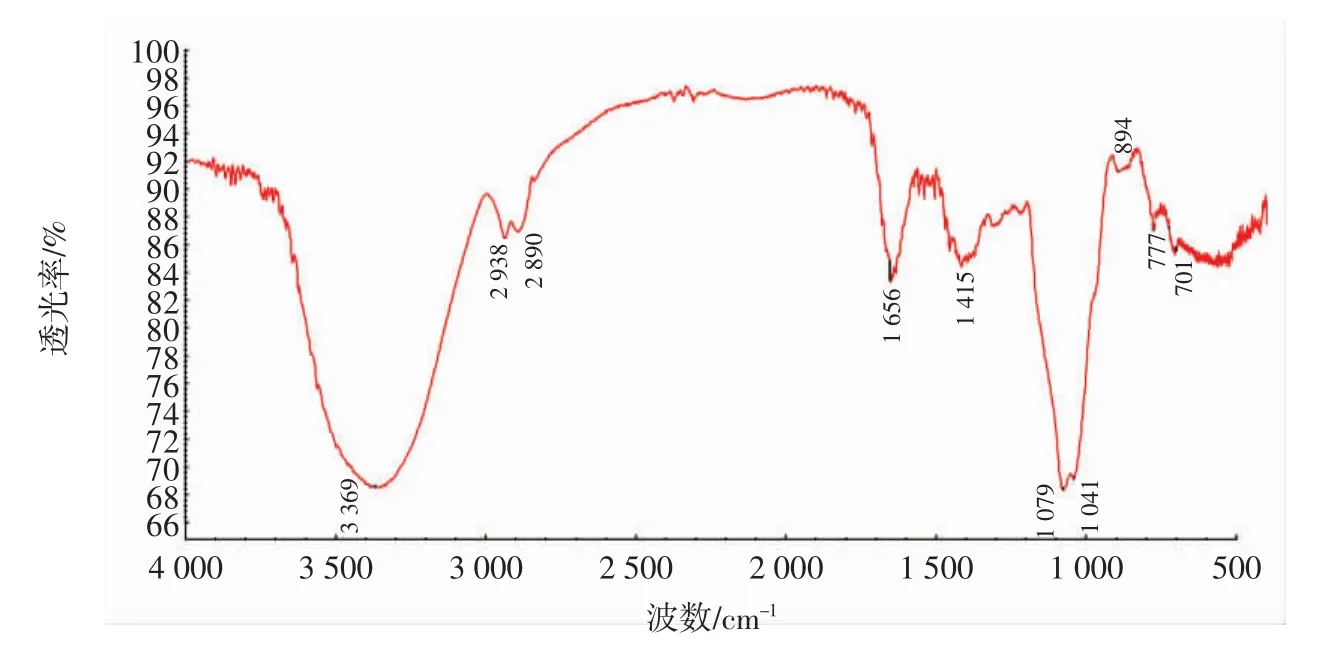
图6 籽瓜多糖SMP-1的红外光谱图Fig.6 Infrared spectrum of SMP-1 of seed melon polysaccharides
由图6可知,多糖类物质的典型特征吸收带在3 600 cm-1~3 200 cm-1处为-OH处的振动吸收峰,该区域的吸收峰,实质上也是糖类表现的特征峰,即3 369 cm-1,可作为糖类相应的特征峰。于1 656 cm-1处有1个吸收峰,归属于C=O的伸缩振动。1 415 cm-1归属于C-O的伸缩振动引起的吸收峰。在1 079 cm-1处有吸收峰为C-O伸缩振动引起的。在1 041 cm-1处吸收峰属于O-H变角振动,该红外存在777 cm-1吸收峰,归属于对称环伸缩振动。896 cm-1存在吸收峰,归属于半乳吡喃糖环的β-端基差向异构的C-H变角振动;但β-端构型推断并不能完全确定,需要做核磁分析进一步确定。
2.7 甲基化分析
单糖残基的类型是由甲基化分析来确定。完全甲基化的籽瓜多糖SMP-1经水解、还原和乙酰化,最后得到部分甲基化的糖醇乙酸酯,然后采用气相色谱质谱联用仪(gas chromatography-mass spectrometry,GCMS)分析检测SMP-1的单糖种类、连接方式以及组成比例,籽瓜多糖SMP-1的甲基化分析见表2。
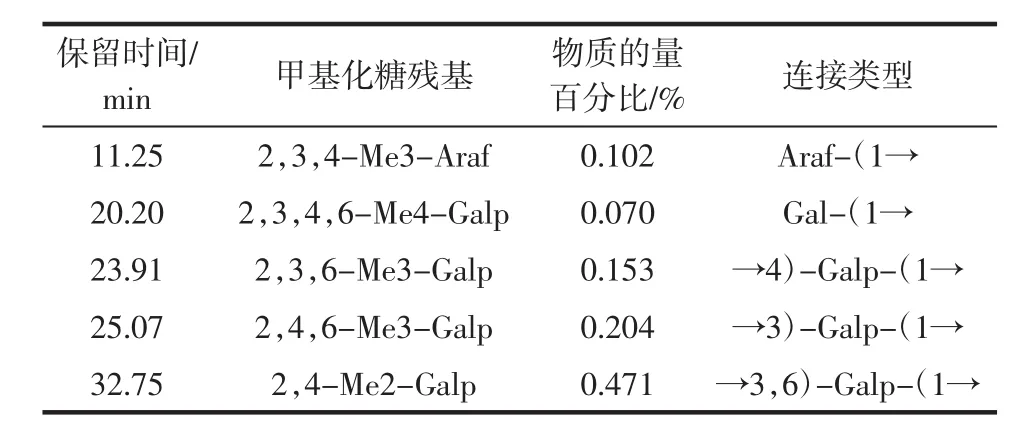
表2 籽瓜多糖SMP-1的甲基化分析Table 2 Methylation analysis of SMP-1 of seed melon polysaccharides
由表2可知,占比较多的是由→3)-Galp-(1→和→3,6)-Galp-(1→组成,糖苷键连接方式较为复杂。
2.8 核磁共振分析
采用核磁共振一维图谱(1H NMR、13C NMR)和二维图谱(COSY、HSQC和HMBC)对籽瓜多糖SMP-1的糖残基组成,以及各糖残基在多糖分子链中的链接方式进行分析,结果见图7和图8。

图8 籽瓜多糖SMP-1的二维NMR图谱Fig.8 Two dimensional NMR spectra of SMP-1 of seed melon polysaccharides
由图7A可知,化学位移介于3.0~5.5。化学位移为3.2~4.0 表示糖环质子信号,基质子峰 5.16、5.01、δ4.56、4.46、4.44的信号峰集中分布在 4.3~5.5。
由图7B可知,核磁碳谱信号化学位移分布于60~120。由碳谱可知,异头碳信号峰为110.62、109.25、105.73、104.79、104.51,异头碳信号区域主要在 93~105。而非异头碳信号峰为 85.22、83.48、82.69、82.43、82.27、81.47、79.04、77.95、77.84、77.53、76.50、76.33、75.87、75.10、74.67、74.33、74.11、74.00、73.20、72.12、71.61、71.23、70.74、70.00、69.85、62.64、62.37、62.10、61.37、非异头碳信号都集中在60~85。根据单糖组成结果,该多糖由阿拉伯糖和半乳糖组成,说明该多糖主要为阿拉伯半乳聚糖。
由图7C可知,在SMP-1的Dept135图谱中,70.72、68.62、62.63、62.41、62.36、62.10 峰为倒峰,表明为 C6的化学位移。70.72向高场迁移,表明存在取代。
籽瓜多糖SMP-1的二维NMR图谱见图8。
由图8A可知,H1-2的信号为4.47/3.57;H2-3的信号为3.57/3.68;H3-4的信号为3.68/4.05;H4-5的信号为4.05/3.87;H5-6a的信号为3.87/3.96,可以推断出H1、H2、H3、H4、H5、H6a 分别为 4.47、3.57、3.68、4.05、3.87、3.96。对应的 C1-C5 为 104.69、71.31、81.5、69.82、74.81、70.76;图8B 显示,异头碳信号为 104.69,HSQC图谱中对应的异头氢信号是4.47。因此,该信号应归属于糖苷键→3,6)-Galp-(1→。
由图8C可知,多糖的糖苷键信号进行归属:糖苷键→4)-β-D-Galp-(1→的异头碳和→3)-β-D-Galp-(1→的H3存在相关峰,可见具有→4)-β-D-Galp-(1→3)-β-D-Galp-(1→;糖苷键→3)-β-D-Galp-(1→的异头碳和→3,6)-β-D-Galp-(1→的 H6b 存在相关峰,可见具有→3)-β-D-Galp-(1→3,6)-β-DGalp-(1→;→3,6)-β-D-Galp-(1→异头氢与自身的→3,6)-β-D-Galp-(1→的 C6 有相关峰,表明存在→3,6)-β-D-Galp-(1→3,6)-β-D-Galp-(1→;糖苷键→3,6)-β-D-Galp-(1→的异头碳 104.69 与→4)-β-D-Galp-(1→的 H4 4.08 有相关信号峰,为→3,6)-β-D-Galp-(1→4)-β-D-Galp-(1→的链接方式。
根据类似规律并结合HMBC图谱和NOESY图谱,对所有糖苷键信号进行归属,结果见表3。
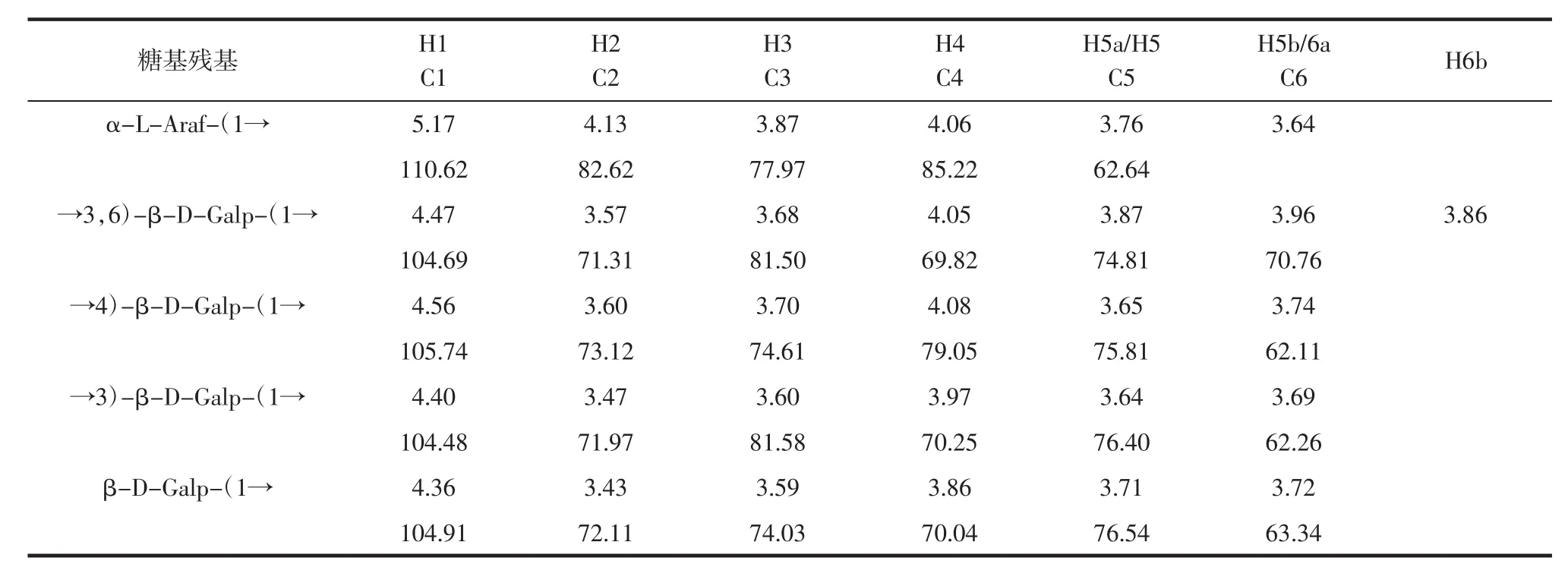
表3 糖苷键信号归属Table 3 Glycoside bond signal assignment
2.9 SMP-1基本结构特征
通过单糖组成、红外光谱、甲基化、核磁共振等结果表明,籽瓜多糖SMP-1的基本结构特征是:SMP-1为1种中性多糖,由半乳糖和阿拉伯糖组成。推断该多糖的主链连接方式为→6)-β-D-Galp-(1→3)-β-D-Galp-(1→的糖苷键,而端基是由 α-L-Araf-(1→通过O-3键连接在主链上。籽瓜多糖SMP-1的一级结构见图9。
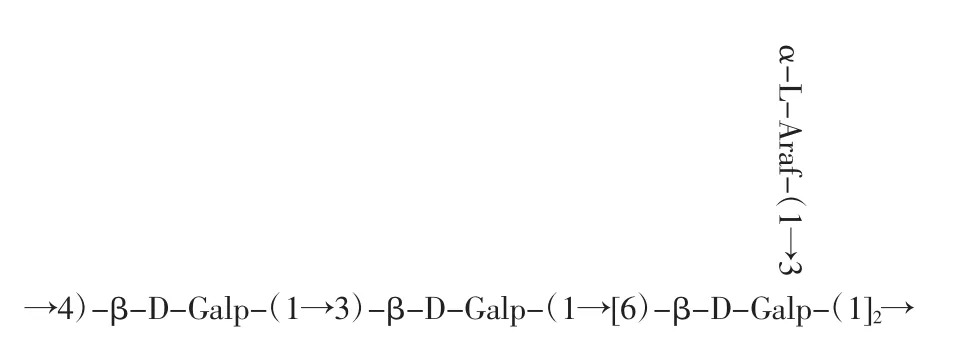
图9 籽瓜多糖SMP-1的一级结构Fig.9 Primary structure of SMP-1 of seed melon polysaccharides
3 讨论与结论
人体肠道菌群存在着多种细菌,是一个复杂的细菌生态系统[15-16]。大量研究表明,多糖能促进益生菌体外生长[17-18]。前期研究发现,废弃籽瓜瓜瓤可以作为获取多糖的重要来源之一[8]。在本试验中,经分离纯化后得到SMP-1、SMP-2和SMP-3 3个多糖组分,它们均能诱导与调节青春双歧杆菌、婴儿双歧杆菌、长双歧杆菌和嗜酸乳杆菌的增殖,且SMP-1的效果优于SMP-2和SMP-3。上述结果表明,籽瓜多糖是一种潜在的益生元且对肠道益生菌具有促进增殖的效果。聚合度会影响糖类对益生菌的增殖作用,聚合度越低其分子量越小,更容易被益生菌利用[19-20]。郝林华等[21]认为多糖对益生菌的促生长作用较明显的为高纯度、β-型糖苷键连接的低聚糖,从而证实益生元作用与糖的结构、纯度等因素有关。因此,基于籽瓜多糖对益生菌生长效应的试验结果,选择SMP-1进一步的结构表征。
多糖自身的生物活性与其分子量、化学组成和结构均有很大的关联[22]。在南瓜中,与杂多糖ATPS-PP-2相比,低分子量的ATPS-PP-1对胰岛细胞具有更好的降血糖活性[23]。较丰富的糖醛酸能改变植物多糖结合物的理化性质和溶解度,具有较高的抗氧化活性[24]。本试验结果表明,SMP-1的分子量为97.40 kDa。其组分由阿拉伯糖和半乳糖构成,所包含的单糖种类较少。与其他植物来源相比,SMP-1的单糖组成与它们存在较大差异,这可能是由品种间差异造成的[25-26]。植物多糖的结构特征,如β-(1)-→3)-在主链中的连接对多糖的生物活动至关重要[27]。由(1→3)和(1→4)-Glc p 糖苷连接的南瓜多糖具有更好的降血糖活性[23]。本试验借助红外光谱扫描、甲基化以及核磁共振波谱分析等多种手段,对SMP-1相应的一级结构予以表征,可推断其结构式为主链连接方式,主要包含的糖苷键为→6)-β-D-Galp-(1→3)-β-D-Galp-(1→,端基是由 α-LAraf-(1→通过O-3键连接在主链上。
综上所述,籽瓜多糖是一种潜在的益生元,可以明显促进青春双歧杆菌、婴儿双歧杆菌、长双歧杆菌和嗜酸乳杆菌的生长。SMP-1构效关系分析表明,籽瓜多糖SMP-1的促生长作用可能与其分子量、单糖组成和糖苷键结构的差异有关。籽瓜多糖SMP-1的主链连接方式为→6)-β-D-Galp-(1→3)-β-D-Galp-(1→的糖苷键,而端基α-L-Araf-(1→通过O-3键连接在主链上。本研究主要分析籽瓜多糖对于益生菌生长的促进效应,为籽瓜多糖的进一步开发利用奠定基础。为了深入了解籽瓜多糖的作用机理,有必要进一步分离纯化籽瓜多糖,并对其高级结构和功能进行详细研究。
