水生植物化感作用抑藻研究进展
2021-08-24于鲁冀彭赵旭范鹏宇李廷梅任昆阳
汤 鹏,于鲁冀,,彭赵旭,范鹏宇,李廷梅,任昆阳
(1. 郑州大学 生态与环境学院,郑州 450001;2. 郑州大学 水利科学与工程学院,郑州 450001;3. 郑州大学 环境政策规划评价研究中心,郑州 450003)
近年来,由于城市化、工业化发展加快,人类生产和生活过程中产生大量废水,部分废水未经完全处理而被排放到天然水体中,导致水体中氮磷等营养元素含量剧增,水体富营养化现象严重,藻类水华频繁暴发[1-2]。目前,如何抑制藻类水华暴发已经成为一个世界性难题。传统的物理方法——遮光、曝气、打捞等不仅耗时耗力且成本过高,处理后依然存在藻类二次暴发的风险;化学投药法虽然能达到快速杀藻的目的,但是会造成二次污染,只能作为应急措施采用;生物滤食和投放微生物作为生物法,存在不可控的风险,在藻类已经大范围暴发的前提下处理效果不理想[3]。作为原材料的水生植物来源广泛,价格低廉;另外不投加化学物质,不产生二次污染,是一种生态友好的方法,现在愈来愈受到研究者们的关注。
相较于陆生植物,水生植物生长迅速、次生代谢产物更加丰富,在自然条件下与藻类距离更近可有效减少水生植物的代谢产物在中途的损失,在实际的抑藻应用中潜力更大。研究水生植物对藻类的化感抑制作用,理解化感作用机理,不仅能指导水华的预防和控制,而且可以为生态不完全水体的功能群恢复提供物种选择和调控。本文总结了水生植物的化感作用及其作用物质、机理,并展望了今后水生植物抑藻的研究方向。
1 水生植物化感抑藻作用
1937年,H. Molisch首次提出了植物化感作用(Allelopathy)这一概念,到1984年Rice在《Allelopathy》第二版中明确了植物化感作用的定义:植物通过将某种化学物质释放到环境中从而直接或间接地对其他植物造成有利或者不利的影响[4]。植物化感作用的研究与应用最初主要集中于农业、林业、环境生物治理等领域,直到20世纪末期,关于水生植物化感抑藻的研究日益增多。研究之初,针对水生植物是否通过化感作用来抑制藻类存在许多争论,水生植物对藻类的抑制效果主要来源4种情况:(1)水生植物与藻类的营养竞争导致;(2)水生植物遮蔽藻类生长所需的阳光导致;(3)水生植物根系的微生物作用导致;(4)水生植物的化感作用导致。目前大量研究已证实水生植物对藻类的化感抑制作用,如Nakai等[5]通过共同培养实验,在排除营养竞争、光照遮蔽及微生物作用的可能后,证实其受试水生植物金鱼藻和穗状狐尾藻是通过连续释放化感物质对席藻产生化感抑制作用;而这种化感抑制效应在野外模拟试验中也有显著影响。姚远等[6]通过控制光照和营养盐浓度进行微宇宙模拟试验,发现苦草、金鱼藻和穗花狐尾藻皆能通过化感作用特异性抑制藻类生长从而调控杭州西湖湖西湿地的藻类群落结构。国内外已经报道了很多具有化感抑藻作用的水生植物,主要包括挺水植物、浮水植物、沉水植物等,常见的具有化感抑藻作用的水生植物如表1所示。

表1 常见具有化感抑藻作用的水生植物Table 1 Common allelopathic algae-inhibiting aquatic plants
2 水生植物化感抑藻物质
化感物质是水生植物化感抑藻作用的关键,水生植物的次生代谢产物——化感物质必须先释放到水体中,通过水环境的传送作用于目标藻类从而起到抑藻的效果。迄今为止,已知的化感物质主要分为14类:水溶性有机酸、直链醇、脂肪族醛和酮;简单不饱和内酯;长链脂肪族和多炔;萘醌,蒽醌和复合醌;简单酚,苯甲酸及其衍生物;肉桂酸及其衍生物;香豆素类;类黄酮;单宁;类萜和甾类化合物;氨基酸和多肽;生物碱和氰醇,硫化物和芥子油苷;嘌呤和核苷[4]。水生植物生长代谢过程中通常会释放多种化感物质,而从水生植物种植水和浸提液中检测到化感物质主要为类萜化合物、简单酚、苯甲酸及肉桂酸衍生物等[22-25]。
水生植物日常代谢释放到水体中的化感物质浓度低,在100 FW/L的种植密度下单种化感物质含量一般不超过100 μg/L,而大部分化感物质的有效抑藻浓度往往要高于5 mg/L[24],即水体中监测到的化感物质浓度一般低于其抑藻要达到的浓度。为了使水生植物化感抑藻更加高效便捷,这就需要获取高浓度的化感物质,通常通过萃取分离等手段从水生植物体内进行提取。如高云霓等[25]为研究苦草释放的化感物质对铜绿微囊藻的抑制作用,通过广谱性固相萃取小柱富集培养3 d的苦草种植水;荆晓玲等[26]则采用乙醇萃取芦苇干物质,再经过过滤分馏等步骤从芦苇中分离得到具有强抑藻能力的化感物质2-甲基乙酰乙酸乙酯(EMA),并发现EMA能选择性地抑制铜绿微囊藻和蛋白核小球藻的生长,而对普通小球藻的生长并没有明显的抑制效果。另外不少研究也发现高浓度的化感物质才对藻类生长起到抑制作用,浓度低时反而促进藻类生长[8,25-26],这种现象叫作“低促高抑”。
化感物质对藻类的抑制效果除了具有选择性、低促高抑等特点外,它们之间还存在协同、相加和拮抗作用,这一点在前人的研究和综述中已得到证实[24-25]。
3 水生植物化感抑藻作用机制
目前关于水生植物化感抑藻作用机制的研究多集中于细胞水平,主要关于影响藻类的光合系统、细胞膜通透性、酶活性等方面,而涉及基因水平的研究较少。
3.1 细胞水平
3.1.1 对光合系统的影响
光合作用是藻类生长必不可少的条件之一,水生植物释放出的化感物质可以通过破坏藻类的光合系统,从而影响藻类的生理状态进而达到抑藻的目的。大量研究表明化感物质对藻类光合系统的影响主要分为降低藻类细胞中叶绿素含量、阻断电子传递两个方面。
降低藻类细胞中叶绿素的含量,其机理是减弱光合能力。Gao等[27]发现化感物质壬基酸可以降低小球藻和月牙藻细胞叶绿素含量,且小球藻表现得更为敏感。朱俊英等[28]利用穗花狐尾藻和铜绿微囊藻的共培养体系,发现铜绿微囊藻的光合作用能力受到化感物质的显著抑制,在处理第3天时,铜绿微囊藻叶绿素含量及叶绿素荧光参数明显降低。相较于叶绿素而言,藻胆蛋白更易受到化感物质的抑制。吴程等[29]发现藻胆蛋白是粉绿狐尾藻分泌的化感物质作用于铜绿微囊藻的靶位点,粉绿狐尾藻通过释放化感物质导致藻细胞中的叶绿素a和藻胆蛋白含量下降,且藻胆蛋白表现得更敏感受损。
阻断电子的传递,其机理是破坏光合系统Ⅱ。Zhu等[19]的研究表明作为化感物质之一的焦性没食子酸能够阻断电子在铜绿微囊藻光合系统Ⅱ上的传递,抑制光合系统Ⅱ从而影响铜绿微囊藻的正常生长。另外刘瑞等[30]也发现采用高浓度N-苯基-2-萘胺处理拟柱胞藻,其藻细胞光合系统遭到损伤,电子在光合系统Ⅱ的传递受阻,抑制了整个细胞的光合作用。
3.1.2 对细胞膜的影响
细胞膜是藻细胞重要的结构和功能基本单位,化感物质可以通过影响藻类细胞膜的结构,破坏其通透性,导致细胞内物质大量渗出,从而抑制藻类生长。董颖娜等[8]发现香蒲水浸提液能明显增大铜绿微囊藻细胞膜的通透性,相较于对照组而言,采用10.00 g/L香蒲水浸提液处理后的铜绿微囊藻细胞膜透性是其1.80倍。而胞内外离子的交换速率会随着细胞膜通透性的增强而加快。荆晓玲等[26]发现鄱阳湖本土水生植物菰的浸泡液可以改变铜绿微囊藻细胞膜通透性,离子通道开放紊乱使渗透压改变,藻细胞直径逐渐缩短,体积减小,藻细胞程序性死亡。别聪聪等[31]发现在化感物质邻苯二甲酸二丁酯(DBP)的作用下,短裸甲藻细胞膜通透性发生变化,24 h后靠近细胞膜处出现了大量的小体积液泡和明显的淀粉粒;同时细胞膜出现大量的突起,使藻细胞生理状态受到损伤。随着藻细胞内离子的大量渗出,水体中各种物质进入细胞内,而细胞内含物渗出丢失,各种细胞器受到不可逆损伤,加速了藻细胞死亡。如杨小杰等[14]发现在凤眼莲和铜绿微囊藻共同培养体系中,仅几天的培养藻细胞就出现了萎缩现象,类囊体发生分解,细胞结构特别是叶绿体、线粒体产生显著损伤,导致细胞生长所需的能量和有机物供给不足,细胞代谢活性降低,生长受到抑制。
3.1.3 对酶活性的影响
酶是生物体不可缺少的物质,藻细胞的新陈代谢过程离不开酶的催化作用。正常情况下藻细胞内的活性氧处于动态平衡状态,水生植物通过释放影响酶活性的化感物质,改变藻细胞的生理状态,引起细胞内活性氧积累,使藻细胞的抗氧化系统受到破坏,从而抑制其生长。施军琼等[32]发现棕鞭藻滤液中存在某些化感物质能引起铜绿微囊藻丙二醛(MDA)和过氧化氢酶(CAT)的含量显著增加,诱导铜绿微囊藻细胞的膜脂过氧化,使藻细胞受到氧化损伤。凤眼莲根系分泌物可以在短时间内使栅藻的超氧化物歧化酶(SOD)活性显著下降,这时活性氧的产生和去除失去了平衡,活性氧的过量积累超过了藻细胞的耐受极限,藻体死亡[14]。
化感物质能影响藻细胞的酶活性,但由于酶的特性不同,化感物质在提高某些酶活性的同时又能抑制另外一些酶的活性。马浩天等[33]发现斜生栅藻细胞在遭到化感抑制的情况下,POD、CAT含量会升高用以对抗活性氧的迸发,而SOD活力下降,可能是SOD合成结构遭到破坏。
除抗氧化物酶活性外,腺嘌呤核苷三磷酸(ATP)酶活性的大小也是藻细胞新陈代谢的重要指标,其活性的减弱会导致藻细胞对钙、镁等矿质元素的吸收等抑制,加剧藻细胞的死亡。汪丽等[34]发现荇菜种植水可以使铜绿微囊藻的ATP酶活性一直保持在低水平状态,培养第9天时其酶活性仅为对照组的16.80%。
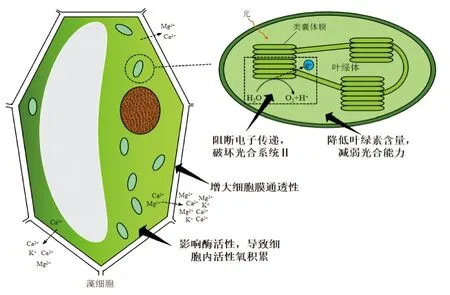
图1 水生植物化感抑藻机制Figure 1 Mechanism of allelochemical inhibition of algae in aquatic plants
3.2 基因水平
水生植物化感抑藻在基因水平方面的研究起步较晚,相关研究较少,主要集中在藻细胞的基因表达水平以及脱氧核糖核苷酸(DNA)的翻译和转录两方面。
化感物质可以增加或降低某些基因表达量,破坏藻细胞正常的生理状态。Lu等[35]发现表没食子儿茶素-3-没食子酸酯(ECGC)诱使铜绿微囊藻细胞的光合系统Ⅰ和光合系统Ⅱ的基因表达量降低,光合系统受到损伤;同时藻胆素降解蛋白的基因表达量明显上升,加速藻胆素的降低,导致捕捉和吸收光的能力下降,从而进一步降低光合作用的效率,达到抑藻的效果。Wang等[36]发现氧化及DNA损伤是化感物质焦性没食子酸对铜绿微囊藻TY001的主要化感抑制渠道,铜绿微囊藻TY001细胞的DNA修复基因(recA和gyrB)上调以及胁迫反应基因(prx、ftsH、grpE和fabZ)的表达水平明显降低,引起抗氧化酶活性上升。化感物质还可以阻碍DNA的翻译和转录,导致蛋白质的合成受阻,其对藻细胞的生理状态影响更为直接。Shao等[37]探究了焦性没食子酸对铜绿微囊藻的胁迫反应基因(prx)、藻毒素合成相关基因(mcyB)以及DNA修复基因(recA、grpE、fabZ)的表达情况,结果表明蛋白质是焦性没食子酸抑藻的一个重要靶位点,焦性没食子酸通过影响蛋白质的合成从而使铜绿微囊藻细胞受到损伤。
4 结论
总体来说,水生植物化感抑藻目前已经进行了大量的研究,在化感物质的分离、鉴定及基于细胞水平的机理研究方面已取得了一定的进展。在水体富营养化治理及水生态修复的迫切需求下,水生植物化感抑藻具有一定的研究价值和应用价值。在今后的水生植物化感抑藻研究中,可以从几个方面开展:(1)目前对水生植物化感抑藻机理的研究主要还停留在细胞水平,而关于化感物质作用于藻细胞基因的靶位点及藻基因是如何变化的没有进行深入的研究。建议从基因水平出发,进一步揭示水生植物抑藻的机理。(2)自然状态下水生植物生长代谢所分泌的化感物质含量较少,释放到水体中后又受到水中生物和非生物因子的影响,故当藻类大量暴发时难以起到很好的抑制效果。因此,具有高效抑藻作用的化感物质的人工提取或合成将是未来研究的重点。(3)目前的研究普遍处于实验室水平,应用于工程的实际案例较少,而天然水体中情况复杂多变,实验室的效果不能等同于野外,在今后的研究中应优先开展一些野外模拟试验,以期为实际项目应用做出重要参考。
