纤毛转运蛋白140谱系示踪小鼠模型构建
2021-07-15陈雨贝
陈雨贝, 孙 瑶
(上海牙组织修复与再生工程技术研究中心,同济大学口腔医学院,同济大学附属口腔医院口腔种植科,上海 200072)
细胞纤毛是一种突出于细胞表面的细胞器,几乎存在于所有哺乳动物的细胞表面[1]。细胞纤毛直径约200~300 nm,长度约几微米。它主要由三部分构成:延续于细胞膜特化的纤毛膜;由中心体和转换纤维组成的纤毛基底部;延续于中心粒的微管束构成的纤毛轴丝[2-3]。细胞纤毛可以通过介导Hedgehog、Wnt、PDGF等信号通路影响组织的发育及细胞功能[4]。这些信号是由细胞纤毛转运复合体纤毛转运蛋白(IFT)来传递的。纤毛转运复合体分为IFT-A和IFT-B。IFT-B由14种IFT组成,包括IFT20、IFT22、IFT25、IFT27、IFT46、IFT52、IFT54、IFT57、IFT70、IFT74/IFT72、IFT80、IFT81、IFT88和IFT172,这些蛋白负责细胞纤毛的正向转运。IFT-A由6种IFT组成,包括IFT144、IFT140、IFT139、IFT122、IFT121和IFT43,其中IFT144、IFT140和IFT122组成细胞纤毛的“核心”,IFT139、IFT121和IFT43组成“外围”,负责细胞纤毛的逆向转运[5]。IFT的突变会导致纤毛形态和功能的异常,从而引起多种纤毛病,主要表现为四肢短小、胸部轻度狭窄、失明、肾衰竭、多囊肾、视网膜变性等,并且会导致一系列骨发育不良,甚至由于骨骼发育障碍而导致出生后即刻死亡[6]。因此,根据纤毛病的骨骼表型,多种综合征被归类为短肋骨-多指畸形组,包括Verma-Naumoff综合 征、Majewski综 合 征、Jeune综 合 征 及Ellis-van Creveld综合征[7]。例如,Jeune综合征表现为短肋骨、短长骨、股骨弯曲及干骺端不规则[8]。
IFT140是IFT-A的一种核心蛋白,在维持IFT-A的稳定性中发挥关键作用。IFT140缺失会破坏IFT-A复合体,导致多种纤毛病。Opitz trigonocephaly C综合征、美因茨-萨尔迪诺综合征、Jeune综合征都已被确定为IFT140突变引起的纤毛病[9]。IFT140突变会导致骨发育不良、视网膜营养不良、肾衰竭等[10-11]。因此,了解IFT140在生长发育中起到的作用对理解纤毛病至关重要。
传统的免疫组织化学染色、免疫荧光染色等只能利用抗体来观察某一特定时间点IFT140的表达,并且实验结果受限于抗体的特异性。细胞谱系示踪技术不需要抗体,其利用Cre/loxP系统标记表达IFT140的细胞,并对其后代的增殖、迁移及分化活动进行追踪和动态观察,结果可靠,且可以获得更多的细胞迁移和分化信息。因此,建立IFT140-CreER;R26RtdTomato示踪小鼠模型对于研究IFT140和细胞纤毛的体内功能非常重要。
1 材料和方法
1.1 实验动物
本实验先设计能够被tamoxifen诱导Cre酶表达的IFT140-CreER小鼠,由赛业生物科技有限责任公司构建(图1A),R26RtdTomato小鼠购买于赛业生物科技有限责任公司。对2种小鼠交配后的子代进行基因型鉴定后,将其用于后续实验。基因鉴定的引物 序 列 为IFT140-CreER-F:5′-CTAATGGTTCCAGTCTGCAGGCCC-3′;IFT140-CreER-R:5′-CAATAAGACCAGGCACATACCATC-3′。小鼠均饲养于同济大学附属口腔医院SPF级动物室内,给予12 h光照和12 h无光饲养环境,室内温度保持在25 °C,相对湿度为55%,给予自由饮水与饮食。所有动物实验经同济大学伦理委员会批准(批准号:TJLAC-017-027)。
1.2 小鼠体内示踪
对IFT140-CreER;R26RtdTomato小鼠行tamoxifen腹腔注射,剂量为0.15 mg/g[12]。IFT140-CreER经tamoxifen激活后,雌激素受体 (estrogen receptor,ER)变为活性状态,暴露出ER的核定位信号,诱导Cre重组酶进入核中,行使其剪切功能[13-14],切除tomato基因序列前2个loxP之间的stop序列。如图1B所示,此时表达IFT140的细胞及其子代可以发出红色的荧光。于出生后3 d(P3)的IFT140-CreER;R26RtdTomato小鼠腹腔内注射tamoxifen,并在5 d后收样(P3+5),观察不同组织中IFT140阳性细胞的分布情况。于出生后4 d(P4)的IFT140-CreER;R26RtdTomato小鼠腹腔内注射tamoxifen,并在注射后3、7、14 d时收样,观察在股骨生长发育过程中,IFT140阳性细胞的分布与转归。
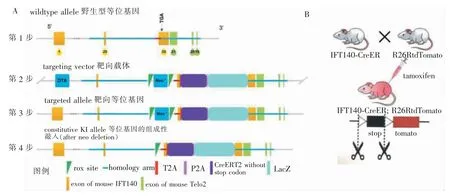
图1 IFT140-CreER;R26RtdTomato示踪小鼠模型构建Figure 1 Generation of IFT140-CreER;R26RtdTomato mouse model
1.3 免疫荧光染色
将IFT140-CreER;R26RtdTomato小鼠处死后用4%多聚甲醛固定24 h,将小鼠的舌、大脑、下颌骨、股骨分离,硬组织用10%乙二胺四乙酸(ethylene diamine tetraacetic acid,EDTA)脱钙1周;用15%、30%蔗糖脱水,用OCT(optimal cutting temperature compound)包埋剂对组织进行包埋,切成10 μm切片。行4′,6-二脒基-2-苯基吲哚 (4′,6-diamidino-2-phenylindole,DAPI)法染色,置荧光显微镜(尼康公司,日本)下观察示踪结果。
2 结果
2.1 IFT140阳性细胞示踪小鼠模型的构建与鉴定
通过将IFT140-CreER小鼠与R26RtdTomato小鼠交配,得到IFT140-CreER;R26RtdTomato小鼠,对其进行基因型鉴定,结果显示,IFT140-CreER目标条带为350 bp,野生型条带为267 bp(图2)。通过注射tamoxifen,得到IFT140阳性细胞示踪小鼠模型。

图2 IFT140-CreER小鼠基因型鉴定Figure 2 Genotype identification of IFT140-CreER mice
2.2 IFT140-CreER;R26RtdTomato小 鼠 体 内 示 踪IFT140的验证
对出生后3 d的IFT140-CreER;R26RtdTomato小鼠进行tamoxifen腹腔注射,标记此时表达IFT140的细胞,并在5 d后收样,发现在纤毛丰富的组织,如磨牙、切牙、毛囊、鼻腔、大脑、舌中有IFT140阳性细胞的特异性分布,说明模型构建成功(图3)。因此,通过IFT140-CreER;R26RtdTomato示踪小鼠模型来观察IFT140阳性细胞在小鼠组织中的增殖、迁移和谱系分化是可行的。

图3 IFT140阳性细胞在体内的分布Figure 3 Distribution of IFT140 positive cells in vivo
2.3 IFT140阳性细胞在股骨中的表达
为了动态探究IFT140阳性细胞在一次骨化中心发育过程中的分布变化,选取出生后4 d的小鼠(P4),对其腹腔注射tamoxifen,标记此时表达IFT140的细胞,并在注射后3 d(P4+3)、7 d(P4+7)、14 d(P4+14)时收样(图4A)。小鼠股骨脱钙后,免疫荧光观察发现,P4标记的IFT140阳性细胞在经过3 d的示踪后,少量分布于骨髓腔内及骨小梁表面(图4B、4E);在经过7 d的示踪后,骨髓腔内及骨小梁表面的IFT140阳性细胞呈增加趋势(图4C、4F);在经过14 d的迁移分化后,骨小梁表面的IFT140阳性信号大量增加,骨皮质内外膜处也呈现出明显的IFT140阳性信号(图4D、4G)。利用Image J软件对阳性细胞面积进行定量,结果显示,在骨发育的高峰时期,IFT140阳性细胞面积呈明显的上升趋势(图4H)。

图4 在股骨发育早期IFT140阳性细胞的分布Figure 4 Distribution of IFT140 positive cells in early femur development
3 讨论
19世纪,Charles O.Whitman和他的同事及后辈开创了细胞谱系示踪技术,并通过光学显微镜发现了无脊椎动物的早期分裂。而发育从最早的分裂开始,个体细胞的命运在发育时是不同的,每个细胞都会产生在后期发育中有特定作用的细胞[15]。同时,它们认识到新的细胞是来源于已经存在的细胞,而不是自发产生的。目前已经发展出多种方式来对某一特定的细胞进行谱系示踪,包括转染、转基因与基因打靶及内源性标记的活体示踪[16]。自20世纪90年代以来,基因工具的快速发展彻底改变了谱系示踪的方法。通过在细胞中引入荧光蛋白或酶(如半乳糖苷酶和碱性磷酸酶)等遗传标记,各种各样的细胞谱系都可以被示踪。Cre/LoxP重组酶系统是目前应用最为广泛的谱系示踪方法,在荧光蛋白的报告基因前插入2个LoxP位点及位点中的停止序列,Cre重组酶可以识别特异的LoxP位点;当Cre重组酶被激活时,停止序列被切除,报告基因可以在所有表达Cre重组酶的细胞及它们的后代中表达。Cre重组酶还可以与突变的ER结合,形成定位于胞质中的融合蛋白(Cre-ER),只有在给予tamoxifen后,Cre重组酶才能进入细胞核发挥作用,从而可以在特定的时间去激活某一类型的细胞,并使报告基因表达[17]。
已经有许多学者利用细胞谱系示踪技术研究某一特定分子在小鼠生长发育中的作用。例如,利用PDGFRβ-P2A-CreER小鼠探究周细胞在血管生成中的作用[18];利用Gli1-CreER小鼠发现了Gli1标记着负责骨形成与骨修复的间充质祖细胞[12];利用Osx-CreER小鼠发现了Osx标记着骨发育中的祖细胞[19]。因此,细胞谱系示踪技术是发现某种关键基因阳性细胞体内功能的重要实验技术。上述实验已经证实,利用IFT140-CreER;R26RtdTomato小鼠可以示踪IFT140阳性细胞在体内的分布。小鼠IFT140突变会导致肋骨侧支、骨化结节[8];而临床研究表明,人类IFT140突变会导致胸廓狭窄、骨骺狭窄、股骨颈变宽[20]。IFT140突变严重影响了骨发育,骨发育需要IFT140的参与。本课题组前期研究发现,IFT140在骨组织中有大量表达,并且年轻小鼠骨组织中的IFT140表达量明显高于老年小鼠[21]。但是上述研究只是静态描述某个时间点的IFT140在成骨细胞中的表达,还不能全面阐述IFT140阳性细胞在体内的分化特征和潜在功能。
因此,本研究构建了IFT140阳性细胞谱系示踪小鼠模型,观察IFT140阳性细胞在小鼠体内的分布和转归。结果显示,IFT140阳性细胞在纤毛富集的器官尤其是在牙齿、骨髓腔、骨小梁处均有较明显的分布,表明体内示踪IFT140阳性细胞的小鼠模型构建成功。利用该示踪小鼠模型不但可以观察IFT140阳性细胞的分布位置,还可以观察IFT140阳性细胞的体内转归。通过对小鼠股骨发育不同时间点的示踪,本课题组发现,在骨发育阶段,IFT140阳性细胞的数目随着发育逐渐增多,在骨小梁处的增加越明显,提示IFT140阳性细胞的成骨分化倾向较明显,可能对于骨发育具有重要意义。未来将通过更长时间的示踪、单细胞分析及免疫荧光染色等技术,结合成骨细胞和成牙本质细胞的特异性染色,探究IFT140阳性细胞在骨组织、牙齿生长发育及疾病发生过程中所起的作用,进而研究IFT140突变导致骨组织和牙齿发育不良的机制。该示踪小鼠模型的建立有利于开展细胞纤毛调节骨骼发育、骨骼稳态维持,以及纤毛相关疾病的发生、发展等的相关研究。
