陆地棉纤维发育相关基因GhEXPs的分析及表达研究
2021-06-19吕丽敏左东云王省芬张友平程海亮王巧连宋国立马峙英
吕丽敏,左东云,王省芬,张友平,程海亮,王巧连,宋国立,4*,马峙英*
(1.河北农业大学/棉花生物学国家重点实验室河北基地,河北 保定071001;2.中国农业科学院棉花研究所/棉花生物学国家重点实验室,河南 安阳455000;3.河北农业大学/教育部华北作物种质资源研究与利用重点实验室,河北 保定071001;4.郑州大学/棉花生物学国家重点实验室郑州基地,郑州450001)
扩展蛋白(Expansin)是一类重要的细胞壁蛋白,在酸性条件下它能松弛细胞壁,促使细胞增大[1-2]。扩展蛋白在细菌、真菌和高等植物中以多基因家族的形式广泛存在,在促进细胞伸长、茎尖和根伸长、叶片的形态建成、花和果实的发育、胚胎发生、花粉发育和花粉管生长等过程中发挥着重要作用[3-6]。McQueen-Mason团队最早从黄瓜幼苗中分离出有活性的扩展蛋白,研究表明该蛋白在酸性、离体条件下能够促进下胚轴细胞的伸长[7]。
近年来,随着分子生物学的快速进步和高通量测序技术的发展,科学家从植物和微生物中鉴定了多个物种的扩展蛋白基因家族。通过对拟南芥[9]、大豆[10]、水稻[9]、大白菜[11]、杨树[12]、烟草[13]等植物扩展蛋白的研究发现,植物类扩展蛋白家族包括4个亚家族,分别为α-扩展蛋白(EXPA)、β-扩展蛋白(EXPB)、类α-扩展蛋白(expansin-like A,EXLA)和类β-扩展蛋白(expansin-like B,EXLB)[8]。该家族蛋白通常包含250~275个氨基酸残基,氮端有包含20~30个氨基酸的信号肽,属于分泌蛋白[3]。扩展蛋白包括2个典型的结构域,分别是位于氮端的DPBB(Double-psi betabarrel)和碳端的CBM 63(Fam ily-63 carbohydrate binding module),其中DPBB是1个六链双“ψ”β-桶形结构,CBM 63是碳水化合物结合模块家族-63结构域[4]。结构分析表明,2个扩展蛋白分子中间夹1个细胞壁多糖形成“三明治”式的结构,该结构是扩展蛋白促使细胞壁发生松弛的作用模型[3]。
棉花纤维是胚珠表皮细胞分化伸长的单细胞结构,它的特殊伸长机制一直是生物学研究的热点,棉花纤维是研究单细胞伸长的模式系统[14]。棉花纤维发育过程分为5个时期,分别是起始期、伸长期、转换期、次生壁合成期和成熟期,各个时期之间存在交互重叠,没有严格的界限[15]。扩展蛋白参与棉花纤维细胞伸长发育的生物学过程已取得了一些进展[16-18],科学家利用棉花纤维的cDNA(Complementary DNA,互补DNA)文库、基因芯片、SSH(Suppression subtractive hybridization,抑制差减杂交)文库等技术从棉花中筛选出在纤维优势表达的扩展蛋白基因[19-20],发现EXPA亚家族的2个基因GhExp1和GhExp2在棉花纤维中高表达[21],后来证明这2个基因是位于棉花A亚组和D亚组的1对同源基因[22]。在陆地棉中过表达GhEXPA8基因能显著促进纤维的伸长[23];序列比对发现GhEXPA8和上述的GhExp2是同一个基因。通过对陆地棉扩展蛋白全基因组分析,采用系统命名法将该基因命名为GhEXPA2,染色体定位分析发现它位于D亚组[22]。此外,从海岛棉A亚组获得了1个扩展蛋白基因GbEXPATR,属于EXPA亚家族,过表达该基因能够使棉纤维变得更长、更细,强度更强。与陆地棉GhEXPA2相比,GbEXPATR缺少CBM 63结构域,这可能与它在海岛棉中的特殊功能有关[16]。近来,对陆地棉扩展蛋白基因家族的研究鉴定了93个GhEXPs基因,发现一些基因在纤维发育中上调表达[22]。这些研究表明,扩展蛋白在纤维发育过程中发挥重要作用。
早期科学家认为扩展蛋白通过减弱多糖和纤维素微纤丝之间的连接作用,从而使细胞壁松弛[2],也可能是通过清除纤维素和多糖之间的氢键使细胞壁松弛[8]。随着研究的不断深入,科学家发现细胞壁中的纤维素微纤丝呈束状分层排列,并把纤维素微纤丝交叉的位置称为“生物力学热点(biomechanical hotspots)”,该区域存在的细胞壁多糖使纤维素微纤丝连接在一起[24]。目前,科学家已经获得了细菌扩展蛋白 EXLX1(Expansin-Like X)和玉米EXPB亚家族的扩展蛋白EXPB1(Zea m1)的晶体结构。研究表明,芳香族氨基酸苯丙氨酸、色氨酸和酪氨酸是扩展蛋白和多糖底物相互结合的关键位点,并且十分保守[25-26]。扩展蛋白晶体结构的解析为我们深入了解其生物学功能和机制奠定了坚实基础。
为进一步阐释棉花纤维伸长的作用机制,解析扩展蛋白在棉花纤维发育过程中发挥的作用。本文从陆地棉转录组数据中筛选了4个在纤维发育过程中优势表达的扩展蛋白基因GhEX PA1d、GhEXPA4b、GhEXPA23b和GhEXPB3a,对这4个GhEXPs基因的进化关系、蛋白结构及表达模式等方面进行了研究。
1 材料与方法
1.1 材料
试验材料陆地棉(Gossypium hirsutumL.)标准系TM-1是来源于中国农业科学院种质资源中期库的资源,本课题组种植于中国农业科学院老所部试验基地(河南省安阳县,36°06′84.44″N,114°49′61.5″E)保存使用,田间常规管理。在盛花期对试验材料进行挂牌,分别于开花当天(0 day post anthesis,0 DPA)、3 DPA、5 DPA、7 DPA、10 DPA、15 DPA、20 DPA和30 DPA的胚珠和纤维进行取样,5 DPA及之前时间点的样品是胚珠和纤维未分离的材料,其余时间点的样品均是从胚珠上剥离的纤维材料。冰上取材后立即放入液氮速冻,于-80℃超低温冰箱保存备用。
1.2 方法
1.2.1 总RNA的提取和c DNA制备
RNA提取按照多糖多酚植物总RNA提取试剂盒(货号:DP441,北京天根生化科技有限公司)说明书及参考相关文献进行操作[22]。
提取的RNA放置-80℃保存备用或直接进行反转录,获得的cDNA用超纯水稀释10倍后,-20℃冰箱保存,用于后面实验。步骤参考Takara公司的PrimeScriptTMII 1st Strand cDNA Synthesis Kit(货号:6210A)的说明书及相关文献[22]。
1.2.2 棉花GhEXPs基因的克隆
从棉花基因组数据库(https://cottonfgd.org/)[27]获得4个GhEXPs的开放阅读框,分别为Ghir_D05G018790.1(GhEXPA1d)、Ghir_A04G009030.1(GhEXPA4b)、Ghir_A05G038370.1(GhEXPA23b)和Ghir_A12G014930.1(Gh-EXPB3a)。设计引物扩增每个基因的开放阅读框(Open read frame,ORF),然后把序列连到pMDTM18-T载体上进行测序验证。基因克隆所用的引物序列见表1。

表1 基因开放阅读框扩增的PCR引物Table 1 The primers for GhEXPs gene open read frame am plification
1.2.3 棉花GhEXPs基因的进化分析及三维结构预测建模
利用MEGA 6.0软件对4个GhEXPs蛋白以及其它物种的扩展蛋白进行多重序列比对并构建系统进化树[28]。构建进化树所用拟南芥蛋白序列来自拟南芥TAIR网站(https://www.arabidopsis.org/),陆地棉扩展蛋白序列来自相关报道[22],其他物种的扩展蛋白序列来自于NCBI数据库,用4个GhEXPs蛋白在NCBI数据库进行BLAST。选择了包括单子叶、双子叶植物以及扩展蛋白不同亚家族成员,包括草莓:FaExp2(AF159563),欧洲甜樱桃:PraExp1(AF297521)、PraExp2(AF297522),黄瓜:CsExp1(U30382)、CsExp2(U30460),番茄:LeExp1(U82123)、LeExp2(AF096776)、LeExp3(AF059487)、LeExp4(AF059488)、LeExp5(AF0-59489)、LeExp8(AF184232),水稻:OsExp1(Y07782)、OsExp4(U85246),独 脚 金:SaExp3(AF291659),百日草:ZeExp3(AF230333),欧洲油菜:BnExp1(AJ000885),烟草:NtExp1(AF04-9350.1),豌豆:PsExp1(X85187),小麦:TaEXPA2(AAS48871.1),玉米:EXPB1_Zea m1(AAO45-608.1)。
将4个GhEXPs蛋白序列去掉信号肽后,利用 在 线 软 件 Phyre2(http://www.sbg.bio.ic.ac.uk/phyre2/htm l/page.cgi?id=index)进行三维结构模型预测[29],然后利用软件PyMOL V2.3.4对蛋白模型进行可视化处理,再用Adobe Illustrator CS3 13.0.0编辑图片标注关键氨基酸信息。
1.2.4 转录表达分析
从0 DPA、3 DPA、5 DPA、7 DPA、10 DPA、15 DPA、20 DPA和30 DPA纤维和胚珠样品中提取的总RNA进行转录组测序。此外,从Cotton FGD(Cotton Functional Genom icsDatabase,https://cottonfgd.org/)[27]数据库中下载了其它组织样品的RNA-Seq数据,样品包括:根、茎、叶、苞叶、花瓣、雄蕊、雌蕊和花托。测序结果的基因表达量用每千个碱基的转录每百万映射读取的片段数(Fragments per kilobase of m illion mapped reads,FPKM)表示。
1.2.5 目的基因的表达分析
通过qRT-PCR(Quantitative real-time polymerase chain reaction,实时定量PCR)检测0 DPA、3 DPA、5 DPA、7 DPA、10 DPA、15 DPA、20 DPA和30 DPA不同时期的棉花胚珠和纤维样品中4个GhEXPs基因的表达情况。以UBQ7(GenBank登录号:AY189972)为内参基因,引物见表2。试验所用试剂盒为Takara公司生产的TB GreenTMPrem ix Ex TaqTMII Kit(货号:RR820A),采用美国ABI的QuantStudioTM5荧光定量PCR仪。采用2-ΔΔCT方法计算基因的相对表达量[30]。利用Excel 2010进行差异显著性分析。
2 结果与分析
2.1 陆地棉4个GhEXPs基因的克隆及进化分析
本研究中4个GhEXPs基因(GhEXPA1d、GhEXPA4b、GhEXPA23b和GhEXPB3a)开放阅读框分别为768 bp、795 bp、798 bp和804 bp。将GhEXPA1d、GhEXPA4b、GhEXPA23b和Gh-EXPB3a与棉属及其他物种的扩展蛋白一起构建系统进化树(图1),发现GhEXPA4b、EXPA1d和GhEXPA23b分别属于EXPA亚家族的第Ⅳ、Ⅴ和Ⅻ亚组,即EXPA-Ⅳ、EXPA-Ⅴ和EXPA-Ⅻ;GhEXPB3a属于EXPB亚家族的第Ⅱ亚组,即EXPB-Ⅱ(图1)。

表2 定量PCR的引物Table 2 The primers for qRT-PCR
2.2 陆地棉4个GhEXPs蛋白的三维建模及关键氨基酸位点分析
目前植物中仅有玉米EXPB1和细菌中的EXLX1两个扩展蛋白解析了晶体结构,位于EXPB1和EXLX1三维结构表面的芳香族氨基酸是它们和细胞壁多糖结合的关键位点。为寻找棉花纤维细胞壁中扩展蛋白的关键氨基酸位点,本研究把陆地棉的4个GhEXPs蛋白与上述2个蛋白进行序列比对(图2),鉴定关键的氨基酸位点。结果发现,在DPBB结构域中,棉花4个扩展蛋白GhEXPs与玉米EXPB1的色氨酸W 26和酪氨酸Y27对应的位点也是2个保守的芳香族氨基酸,其中GhEXPB3a与玉米EXPB1对应位点的氨基酸完全相同,其余3个GhEXPs与W 26位点对应的氨基为苯丙氨酸(F),4个Gh-EXPs与Y27对应的位点没有发生变异。在CMB63结构域中,玉米扩展蛋白EXPB1的关键位点是Y 160和W 194,4个GhEXPs蛋白与Y160对应的位点变异为W或F(GhEXPA23b除外),与W 194对应的位点完全相同,没有发生变异(图2)。同时,4个GhEXPs与微生物扩展蛋白EXLX1也进行了比对,EXLX1中两个连续的色氨酸W 125、W 126是该蛋白的关键氨基酸位点,4个GhEXPs与W 125、W 126对应的位点变异为YF(GhEXPA23b与W 126对应的位点除外)2个芳香族氨基酸(图2)。

图1 4个GhEXPs蛋白与其它植物扩展蛋白的聚类分析Fig.1 Phylogenetic tree of the four GhEXPs and other plant expansins
利用在线软件建立了陆地棉4个扩展蛋白的三维结构模型,参考模型是玉米EXPB1(模板编号:c2hczX)的结构,建模结果如图3所示。GhEXPA1d、GhEXPA4b、GhEXPA23b和Gh-EXPB3a与玉米EXPB1的序列一致性分别为31%、34%、32%和42%,模型可信度均为100%。从三维结构模型的预测结果来看,GhEXPA1d、GhEXPA4b、GhEXPA23b和GhEXPB3a蛋白均包含DPBB和CBM 63两个结构域,其中DPBB是典型的六链双“ψ”β-桶结构,2个结构域上分布有芳香族氨基酸苯丙氨酸、酪氨酸和色氨酸。我们把序列比对得到的关键芳香族氨基酸标注在三维结构图中(图3),在DPBB结构域中,Gh-EXPA1d的F13、Y14,GhEXPA4b的F18、Y19,GhEXPA23b的F22、Y23和GhEXPB3a的W 19、Y20位于结构的表面;而GhEXPA1d的W 183、F150,GhEXPA4b的W 187、F154,GhEXPA23b的W 190,Y155和GhEXPB3a的W 188、Y154位于CBM 63结构域的表面(图3)。

图2 4个GhEXPs与EXLX1和EXPB1的序列比对Fig.2 Multiple sequence alignment of 4 GhEXPs and EXLX1 and EXPB1
从三维结构图可以看出,每个扩展蛋白均有4个芳香族氨基酸分布在同一平面上,GhEXPA1d包括F13、Y14、F150和W 183,GhEXPA4b包括F18、Y19、F154和W 187,GhEXPA23b包括F22、Y23、W 190和Y155,GhEXPB3a包括W 19、Y20、W 154和W 188(图3),但GhEXPA1d、Gh-EXPA4b和GhEXPB3a中与EXLX1的W 125对应的芳香族氨基酸和它们并不在一个平面上(图3)。而这些位于同一平面的保守氨基酸在细胞壁中与多糖结合可能是松弛细胞壁的关键。
2.3 陆地棉中4个GhEXPs基因的组织表达分析
根据转录组测序结果分析4个GhEXPs基因在棉花纤维发育不同时期及其他组织的表达情况(图4)。总体来看,GhEXPA1d、GhEXPA4b、GhEXPA23b和GhEXPB3a在胚珠和纤维中优势表达,在其他组织包括根、茎、叶、苞叶、花瓣、雄蕊、雌蕊和花托表达水平很低。其中GhEXPA1d和GhEXPA23b这两个基因在纤维中特异表达,在其他组织中基本不表达;GhEXPA4b和Gh-EXPB3a在纤维之外的其它组织如叶片、雄蕊、雌蕊等有较低水平的表达。转录组结果初步分析表明,4个GhEXPs是在纤维发育中明显优势表达的基因。

图3 陆地棉4个GhEXPs蛋白的三维结构预测Fig.3 The predicted three-dimensional structures of 4 GhEXPs
2.4 qRT-PCR分析验证4个GhEXPs基因的表达情况
为进一步验证4个陆地棉扩展蛋白基因的表达情况,利用qRT-PCR技术对4个GhEXPs基因的表达进行了验证。从图5可以看出,4个GhEXPs基因在纤维发育不同时期的表达模式和转录测序的结果基本一致(图4和图5)。GhEXPA1d、GhEXPA4b、GhEXPA23b和GhEXPB3a均在纤维发育的起始期和伸长期上调表达。其中GhEXPA1d和GhEXPB3a在开花当天(0 DPA)表达量相对较高,他们分别在开花后10 d和20 d的表达水平急剧下降(图5);GhEXPA4b和Gh-EXPA23b在花后3~7 d表达量高,然后表达量开始下降(图5)。此外,EXPA亚家族的3个基因在纤维中的表达情况也不相同,GhEXPA1d和GhEXPA4b的表达水平明显高于GhEXPA23b。
3 讨论
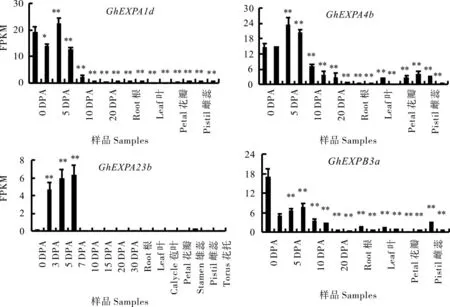
图4 陆地棉4个GhEXPs基因在不同纤维样品和组织中的转录组分析Fig.4 The transcriptome analysis of 4 GhEXPs from fiber and other tissues
本研究鉴定了4个在棉花纤维中特异表达的扩展蛋白基因,其中3个属于EXPA亚家族,1个属于EXPB亚家族,这与陆地棉扩展蛋白基因EXPA亚家族占优势的结果一致[22]。蛋白的一级结构是正确行使功能的基础。在蛋白序列一级结构的DPBB结构域上,对比棉花4个GhEXPs与玉米EXPB1、细菌EXLX1的关键保守位点发现,在这些关键位点也都是芳香族氨基酸,暗示了它们可能与玉米EXPB1和细菌EXLX1具有相似的功能,但个别氨基酸的差异,又凸显了物种之间的不同。
最近利用固态核磁共振(Solid-state nuclear magnetic resonance,ss-NMR)技术揭示了EXPB1蛋白结合的底物是半纤维素葡萄糖醛阿拉伯聚糖(GAX,glucuronoarabinoxylan)[31],点突变试验表明了EXLX1的结合底物主要是纤维六糖[3,32]。在本研究中,根据同源序列比对和建模分析,我们发现陆地棉4个GhEXPs与已报道的解析出晶体结构的玉米扩展蛋白EXPB1的关键氨基酸位点基本相同,并与细菌中已经解析结构的EXLX1蛋白中在关键的芳香族氨基酸位点也有较高的一致性。由此推测本研究发现的棉花扩展蛋白可能具有类似的功能。另有研究发现,细胞壁纤维素在聚合粘连位置形成“生物力学热点”,在这些位点起黏合作用的就是半纤维素成分,扩展蛋白就是通过结合这些半纤维分子把它们从纤维素上解锁从而松弛细胞壁的[24]。棉花纤维细胞壁的半纤维素成分有木葡聚糖、甘露聚糖和葡萄糖醛阿拉伯聚糖(GAX)等[15]。我们推测棉花扩展蛋白也可能是通过与棉花纤维细胞中的半纤维成分结合而使细胞壁发生松弛,从而在内部膨压的驱动下使纤维细胞不断扩展。然而,棉花扩展蛋白在纤维细胞壁内的生物机制以及具体结合的半纤维种类仍需要进一步深入研究。

图5 陆地棉4个GhEXPs基因的qRT-PCR分析Fig.5 qRT-PCR analysis of 4 GhEXPs in upland cotton
扩展蛋白广泛分布在植物和微生物的细胞壁中,通过松弛细胞壁促进细胞的扩展[3,24,33]。研究表明,外加细胞壁蛋白的提取物能够促进细胞伸长[7]。拟南芥根毛特异基因AtEXPA7的RNAi(RNA interference,RNA干扰)试验可明显降低根毛细胞的长度,与野生型对照相比,RNAi株系根毛长度减少了25%~48%[6]。扩展蛋白在参与棉花纤维发育也有诸多报道[16-18],GhEXPA1(即GhExp1[21])在开花后6~28 d的纤维中特异表达且是表达水平高的基因,Xu等[18]研究表明,GhEXPA1和GhRDL1相互作用共同调控纤维的伸长。Li等[16]获得海岛棉扩展蛋白基因GbEXPATR,过表达该基因能明显促进棉花纤维的伸长。扩展蛋白基因具有明显的时空和组织表达特异性。在棉花纤维发育中已经鉴定了位于A亚组和D亚组上的一对同源扩展蛋白基因GhExp1和GhExp2[21],RNA印记实验已经证明二者在纤维发育不同时期高表达,而在根、茎、叶等组织中不表达,超表达GhEXPA8(即GhExp2)基因可以明显促进棉纤维的伸长[23,25]。本研究筛选获得的陆地棉扩展蛋白基因GhEXPA1d、GhEXPA4b、GhEXPA23b和GhEXPB3a也是在棉花纤维中特异表达,在根、茎、叶等组织中表达量低(图4),推测这些扩展蛋白基因在促进纤维伸长发育过程中也可能发挥着重要的作用。
4 结论
本文从陆地棉中获得了4个在纤维中特异表达的扩展蛋白基因,分别是GhEXPA1d、Gh-EXPA4b、GhEXPA23b和GhEXPB3a,前3个基因属于EXPA亚家族,GhEXPB3a属于EXPB亚家族。其编码的蛋白均含有DBPP和CBM 63两个保守结构域。三维结构建模分析发现,芳香族氨基酸苯丙氨酸、色氨酸和酪氨酸分布在扩展蛋白2个结构域表面的4个位点,这些氨基酸处于同一平面上,可能是底物结合的关键位点。4个基因在根、茎、叶、苞叶等组织中表达量低,在棉花纤维组织发育的前期优势表达。本研究为阐明扩展蛋白基因对纤维发育的调控提供了理论依据。
