扁圆吻鲴形态特征及生化遗传特性分析
2021-06-17陈元元林炳明王卫民KianannTan张小东宋晓然魏晋
陈元元 林炳明 王卫民 Kianann Tan 张小东 宋晓然 魏晋


摘要 [目的]從形态特征、染色体核型和乳酸脱氢酶3个层面评估扁圆吻鲴种质资源状况,为丰富鲴亚科鱼类的遗传特性资料,种质评估、种质保护及利用提供理论参考。[方法]采用传统的形态学测量方法对60尾扁圆吻鲴的生物学性状进行描述,采用植物血球凝集素(PHA)和秋水仙素腹腔注射肾细胞直接制片法及同工酶电泳等分子生物学方法,开展扁圆吻鲴的细胞遗传特性及生化遗传特性研究。[结果]扁圆吻鲴体长形,侧扁,腹部圆,肛门前方无腹棱。头小,吻突出。口下位,横裂,下颌有很发达的角质缘。鳃耙短且扁薄,排列很紧密。鳞小,体背部深黑色,腹部银白。背、尾鳍灰黄色,尾鳍边缘黑色,偶鳍基部黄色。鳔分二室,且后室较前室大,前室钝圆,后室末端稍尖且呈锥状;扁圆吻鲴的可数性状依次为背鳍鳍式(D.Ⅲ,6~8)、臀鳍鳍式(A.Ⅱ,6~9)、侧线鳞数(76~89)、鳃耙数(94~103);其体细胞染色体数为2n=48,核型公式为18m+18sm+8st+4t,臂数(NF)为84;其不同采样点的样本眼睛晶状体乳酸脱氢酶(LDH)均有5条同工酶谱带,由LdhA、LdhB两个基因编码。[结论]该研究中扁圆吻鲴的形态特征与已报道的鲴亚科鱼类形态特征存在一定的差异,而体细胞染色体数、核型以及乳酸脱氢酶的表达均与其他鲴亚科鱼类的特征相一致。
关键词 扁圆吻鲴;形态特征;核型;染色体;同工酶
中图分类号 S917.4文献标识码 A
文章编号 0517-6611(2021)02-0077-04
doi:10.3969/j.issn.0517-6611.2021.02.023
开放科学(资源服务)标识码(OSID):
Morphological and Biochemical Genetic Characteristics of Distoechodon compressus
CHEN Yuanyuan1,LIN Bingming2,WANG Weimin3 et al (1.School of Life Sciences,Jianghan University,Wuhan,Hubei 430056;2.Longyan Aquatic Technology Promotion Station,Longyan,Fujian 364000;3. School of Fisheries,Huazhong Agricultural University,Wuhan,Hubei430070)
Abstract [Objective]To evaluate the germplasm resources of Distoechodon compressus from three aspects of morphological characteristics,chromosome karyotype and lactate dehydrogenase,and provide theoretical basis for the enrichment of genetic characteristics,germplasm evaluation,formulation of germplasm standards,germplasm protection and utilization of D.compressus.[Method]The biological characteristics of 60 heads of D.compressus were described by the traditional morphological measurement method.The cell genetic characteristics and biochemical genetic characteristics of D.compressus were studied by the methods of PHA,colchicine and isozyme electrophoresis.[Result]D.compressus had a long body,flat side,round abdomen and no ventral edge in front of anus. It had a small head with a protruding kiss.Its mouth was located at the lower part of the head.The mandible had very developed horny margin.Gill rakes were short,flat and thin,closely arranged.The back of the body was dark black covered with small scales,and the belly was silvery white.Its dorsal and caudal fins were grayish yellow,the caudal fins had black edges,and the base of the even fins was yellow.The swim bladder was divided into two chambers with the smaller blunt and round of anterior chamber.The posterior chamber end was slightly pointed and coneshaped.The four countable characters of D.compressus were dorsal fin type(D.Ⅲ,6-8),gluteal fin type(A.Ⅱ,6-9),lateral scales(10-12),gill rake(94-103).The chromosome number of somatic cells was 2n=48,the karyotype formula was 18m + 18sm + 8st + 4t,and the arm number (NF) was 84.LDH of eye crystal at different sampling points had five isozyme bands,which were encoded by two genes of LdhA and LdhB.[Conclusion]In this study,there were some differences of the morphological characteristics between D.compressus and Xenocyprinae,the number of chromosome,karyotype and expression of LDH in somatic cells are consistent with those of other Xenocyprinae.
Key words Distoechodon compressus;Morphological characteristics;Karyotype;Chromosome;Isoenzyme
扁圆吻鲴(Distoechodon compressus)隶属鲤形目(Cypriniformes)鲤科(Cyprinidae)鲴亚科(Xenocyprininae)圆吻鲴属(Distoechodon),主要分布于我国福建省连城县及台湾省北部等地区。因其食性广、适应性强、疾病少、人工繁殖技术成熟、肉质鲜嫩等特点,扁圆吻鲴是主养和混养的理想种类,在江河、湖泊、水库放流可自然繁殖,是适宜放养增殖的优良品种。自20世纪50年代以来,该品种已在福建省福州、清流、宁化、长汀、永安等地区推廣养殖。此外,扁圆吻鲴属于刮食性鱼类,喜食水体中的着生藻类,在自然水体和养殖池塘中可充分利用饵料资源,有效改善水质和底质环境。为了保护这一优良种质资源,改善水域生态系统,在渔业主管部门的主导下已有不少地区开展了扁圆吻鲴的增殖放流行动。目前有关鲴亚科鱼类的报道主要集中在细鳞斜颌鲴养殖技术与模式[1]、乳酸脱氢酶[2]、线粒体DNA 基因[3]的研究。扁圆吻鲴的研究报道主要集中在生物学特性、人工繁殖[4]、生态分布[5]等方面,而关于扁圆吻鲴种质研究的报道还很缺乏。笔者通过对不同水域、不同生存环境的扁圆吻鲴形态学、染色体和同工酶等基本特性进行研究,旨在为扁圆吻鲴种质资源保护、种质标准和种质鉴定提供有价值的生化遗传参数,从而促进扁圆吻鲴这一品种的开发和利用。
1 材料与方法
1.1 试验材料
试验用鱼于2019年采自福建省连城县,共60尾,体重35.2~154.7 g,体长11.67~20.13 cm。
1.2 形态特征测定
参照GB/T 18654.3—2008《养殖鱼类种质检验第3部分:性状测定》,采用游标卡尺(精确到0.01 mm)对60尾样本鱼进行测定。可量性状包括全长、体长、体高、头长、吻长、眼径、眼间距、尾柄长、尾柄高;可数性状包括背鳍和臀鳍的鳍式、侧线鳞鳞式、鳃耙数。内部构造包括鳔和腹膜。
1.3 染色体制备
随机选取健康的扁圆吻鲴6尾,腹腔注射植物凝集素(4.5 μg/g),24 h待肛门充血发红后即可用于染色体制片。断尾放血后,取出头肾,用0.8%的生理盐水清洗后,用眼科剪剪碎并过目筛绢网,获得头肾细胞悬液。用终浓度0.2 μg/mL的秋水仙素处理50 min后,离心后弃上清,在沉淀物中加入KCl溶液混匀后低渗作用30 min。离心后弃上清,使用新配制的卡诺氏液(甲醇∶冰醋酸=3∶1)预固定2 min后离心,弃上清液,再加入卡诺氏液固定20 min,此过程重复3次。最后,离心后收集沉淀并加入0.5 mL卡诺氏液,制成悬液。采用4℃预冷过的载玻片进行冰冻滴片,空气干燥;预先配制的Giemsa母液按1∶9的比例与磷酸缓冲液(pH 7.8)混合后进行染色,染色晾干后光学显微镜下观察,并在油镜下进行拍照。
1.4 染色体核型分析 参照林义浩[6]的方法,选取来自不同个体、分散良好、形态清晰的中期分裂相进行拍照计数,并计算其相对长度、臂比、染色体的分类与臂数(NF)。参照郭丰等[7]的方法进行染色体分类和分组。染色体臂数:将中部和亚中部着丝点染色体的臂数计为2,亚端部和端部着丝点染色体的臂数计为1。
1.5 组织酶液的制备 参照贺刚等[8]的方法,将扁圆吻鲴断尾放血后取眼睛晶状体,经4 ℃预冷的生理盐水冲洗干净后用滤纸吸干水分,样品称重后放入洗净预冷的匀浆器内,按1∶3(g∶mL)的质量体积比添加双蒸水,冰浴条件下反复研磨成组织匀浆液,4 ℃ 12 000 r/min 离心30 min,吸取上清液,重复2次。离心后获得的上清液用于电泳分析。
1.6 电泳及染色 乳酸脱氢酶同工酶分析采用聚丙烯酰胺梯度凝胶垂直电泳,参照孟彦等[9]的方法,浓缩胶和分离胶浓度分别为4.0%和7.5%,电极缓冲液为pH 8.3的Tris-甘氨酸缓冲液。采用10 mA电流进行预电泳,待其指示剂超过浓缩胶,将电流调节至20 mA,电泳时间为6 h。整个电泳过程在4 ℃条件下进行。
电泳结束后取出凝胶,参照余来宁等[10]的方法,室温避光条件下染色,显带清晰后立刻用蒸馏水冲洗,然后拍照保存。染色液的配制方法如下:1% NAD 3 mL、6% 乳酸钠2 mL、1% NBT 4.8 mL和1% PMS 0.4 mL。
1.7 数据统计与分析 乳酸脱氢酶同工酶以区带数目、染色强度和相对迁移特性为指标进行分析。试验测得的可量性状使用SPSS 19.0软件进行统计与分析。
2 结果与分析
2.1 外部形态特征及可数性状和可量性状
2.1.1 外部形态。体长形,侧扁,腹部圆,肛门前方无腹棱。头小,吻突出。口下位,横裂,下颌有很发达的角质缘。鳃耙短且扁薄,排列很紧密。鳞小,背鳍起点约在体中,与腹棱起点相对,硬刺粗壮而光滑。尾鳍深叉形。体背部深黑色,腹部银白,体侧有10余条黑色斑点组成的纵条纹。背、尾鳍灰黄色,尾鳍边缘黑色,偶鳍基部黄色(图1)。
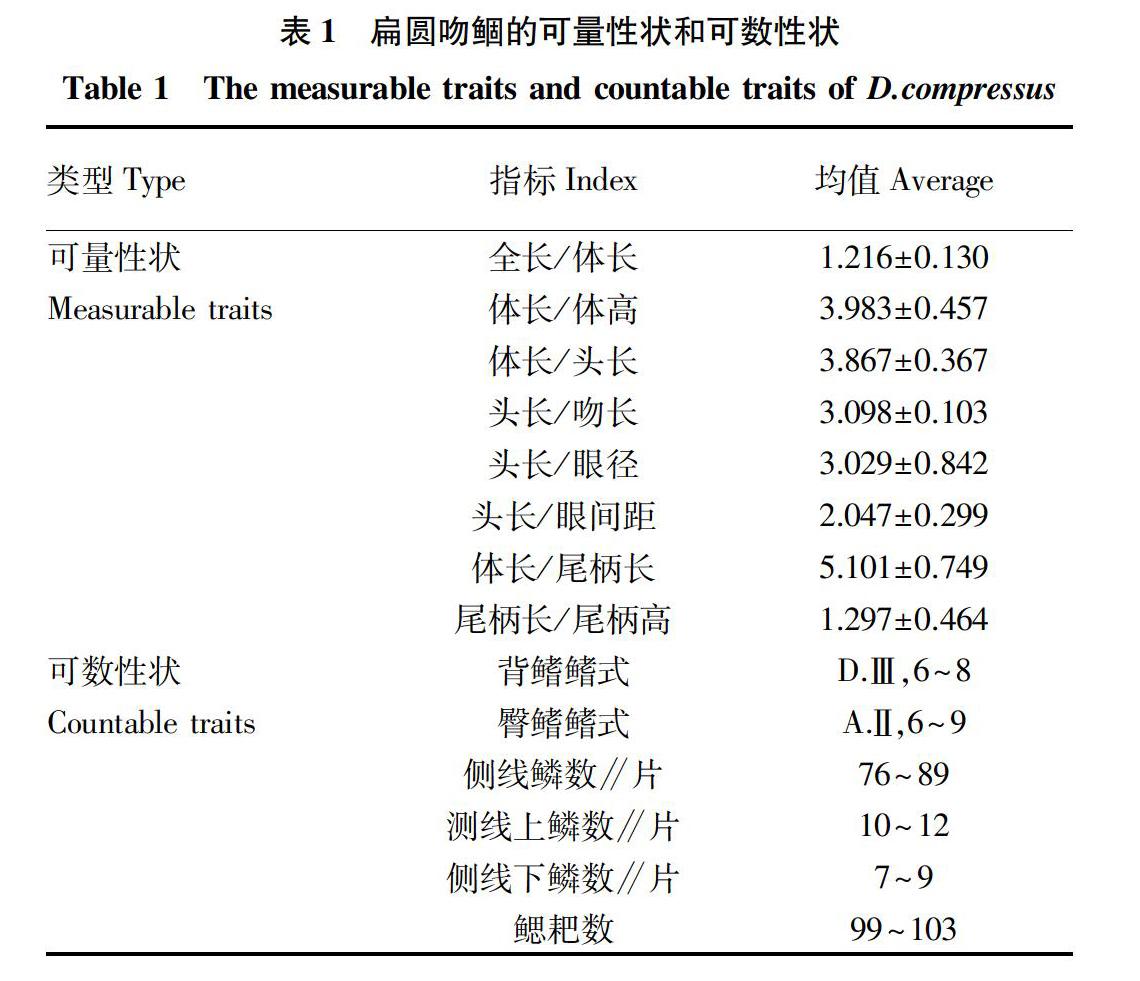
2.1.2 可数性状和可量性状。鳔分二室,且后室较前室大,前室钝圆,后室末端稍尖且呈锥状,腹膜黑色(图2)。扁圆吻鲴的可量性状和可数性状详见表1,均值反映所测数据的集中程度。在可量性状中,主要以体长和头长为参照,测算出吻长和眼间距等头部主要参数与头长的比例,也包括体高、尾柄长和尾柄高等躯干部主要参数与体长的比例。在可数性状中,侧线鳞数的变化范围最大(表1)。
2.2 染色体核型分析 选取无重叠、分散良好、长度适中且着丝粒清楚的分裂相,确定其染色体数目为2n=48(图3)。通过测量染色体相关参数,并使用Photoshop软件将染色体排列好(图4),确定其组型和臂数。根据染色体的着丝点位置,可将扁圆吻鲴染色体核型分为4组:第1组有9对中部着丝点染色体(m),第2组有9对亚中部着丝点染色体(sm),第3组有4对亚端部着丝点染色体(st),第4组有2对端部着丝点染色体(t)。染色体核型为18m+18sm+8st+4t,染色体臂数(NF)为84。
2.3 乳酸脱氢酶(LDH)表达
从图5可以看出,样本鱼眼睛晶状体的LDH酶谱均检测到5条LDH酶带,其中LDH1的表达活性最强,LDH4、LDH5的表达活性相对较弱。对所得酶谱进行扫描分析,结果在样本鱼眼睛晶状体的扫描图中发现5个峰(图6)。
3 讨论
3.1 扁圆吻鲴的形态学比较
该研究检测了60尾扁圆吻鲴样本群体的形态学数据,通过与已报道的鲴亚科鱼类形态特征进行比较,发现该研究结果与前人研究结果
[11]存在一定差异。究其原因,除了与样本量以及样本鱼的规格有关外,种间差异是最大影响因素。通常在形态度量学特征上,鱼类比其他脊椎动物有更大的种群内部和種群之间的变异,且这些形态变异更易受环境的影响[12]。作为独立的生物学单元,不同种群具有不同的适应性进化潜能,其表型与遗传特征也往往存在差异[11]。由于长期的隔离以及对它们各自环境的适应,鱼类不同群体间存在显著的形态差异是可能的。该研究数据可为扁圆吻鲴种质资源保护及种质标准的制定提供基础资料。
3.2 扁圆吻鲴的染色体核型分析
对鱼类的种质评估应结合形态特征、细胞遗传学特征和生化遗传学等特征进行研究。从扁圆吻鲴染色体照片分析得出,样本中同一分裂相中最大的1对染色体的2条同源染色体属于亚中部。这与张林等[13]报道的银鲴等4种鲴科鱼类最大的1对亚中部染色体特征相似。这表明鲴科鱼类中这一对最大染色体的相对长度和臂比可能不会因罗伯逊易位、倒位、缺失或重复等而改变,反映了鲴科鱼类核型的同源性。
该研究发现扁圆吻鲴的染色体数目2n=48,这与李康等[14]对扁圆吻鲴的研究结果相一致。鲴亚科和鳊亚科同属于鲤科鱼类,研究学者对鲤科鱼类染色体的研究报道甚多,秦改晓等[15]认为其染色体的基本二倍数为2n=50和2n=48,变异范围为44~200。洪云汉等[16]研究表明中国鲤科的核型为2n=50。从染色体数目来看,扁圆吻鲴与已报道的圆吻鲴、银鲴、黄尾鲴、细鳞斜颌鲴、四川鲴、方氏鲴等[14]其他鲴亚科鱼类染色体数目一致,均为2n=48,可推测鲴亚科鱼类与其他许多鲤科鱼一样,染色体数目在进化上是保守的。马纲[17]认为某一分类群大多数或绝大多数种类所具有的染色体二倍数,可作为该分类群最基本的核型特征。扁圆吻鲴与已知鲴亚科鱼类[如黄尾鲴(2n=48)、银鲴(2n=48)、细鳞斜颌鲴(2n=48)、逆鱼(2n=48)等]的二倍体数一致。
除了染色体数目外,扁圆吻鲴的核型与圆吻鲴、黄尾鲴和其他鲴亚科鱼类的核型不一致,圆吻鲴、细鳞斜颌鲴和逆鱼的核型均为18m+26sm+4t(NF=92),银鲴核型为20m+26sm+2t(NF=94)[14,17-18],而扁圆吻鲴的染色体核型为18m+18sm+8st+4t(NF=88)。根据现代鱼类演化的理论,染色体结构的重排可以引起NF值的变化,在2n值相同的分类群之间,臂数的变化是从进化上低位类群到高位类群的表现为逐渐升高的趋势,即具有较多端部着丝粒染色体的类型是比较原始的,而出现较多中部和近中部着丝粒染色体的类型是比较进化的[17]。因此,从核型来看,扁圆吻鲴与圆吻鲴、黄尾鲴、细鳞斜颌鲴和逆鱼的进化地位更相近,而银鲴的进化程度更高。上述核型多态性也可能是由于鱼类的不同地理种群,不同学者使用的方法不同,测量染色体的时相不一致以及测量和配组误差所造成的。
3.3 扁圆吻鲴LDH同工酶的比较分析
基因在生物漫长的系统发育过程中也在不断进化与变异。经过突变积累以及自然选择作用改变了其基因调控方式,导致核苷酸序列发生变异,形成另一个基因,然而这些基因最初均来源于一个“祖先基因”。Markert等[19]对鱼类LDH基因的进化进行了详细阐述,通过进一步分析基因进化过程中LDH同工酶的分布和变化,可反映出低等无颌类到高等硬骨鱼类的某些进化性的差异。吴力钊等[20]研究发现白鲢和团头鲂的胚胎发育过程中某些特异性同工酶的开始合成与发育过程中的细胞分化或器官形成时的特殊事件有关。在硬骨鱼类中具有标志性的Ldh-C基因是在鱼的眼睛中发现的,因此笔者以扁圆吻鲴鱼的眼睛晶状体为样本来检测同工酶。
LDH为四聚体酶,鱼类的LDH同工酶最少有3个基因编码,其中A、B基因所编码的酶出现在鱼类的眼睛晶状体中。该研究在9尾扁圆吻鲴的晶状体样本中检测到LdhA、LdhB基因编码的经典5种酶带,可用于种质鉴定。在样本组织中未见LdhC基因的表达,说明扁圆吻鲴LDH同工酶的遗传多样性处于中等地位。Shaklee等[21]认为第3个基因所编码的酶在鲤形目中只存在于肝脏中。同一物种不同组织中LDH同工酶谱存在差异,即具有组织特异性,不同亚基组成的LDH同工酶作用不同,这一特征与该组织所处的生理条件和机能有关[22]。
LDH同工酶数量与动物生活环境含氧量的状况相关。该研究中在扁圆吻鲴的晶状体中检测到5条的经典酶带,数量处于中等,由此推断扁圆吻鲴适宜在含氧量适中、比较稳定的水域环境中生长,这可能与扁圆吻鲴在水体中下层活动生活习性有关。
4 结论通过对扁圆吻鲴的形态特征和遗传学特性的研究发现,扁圆吻鲴形态特征与鲴亚科鱼类的细鳞斜颌鲴存在一定差异。扁圆吻鲴的体细胞染色体数为2n=48,核型公式为18m+18sm+8st+4t;染色体臂数(NF)为84。其晶状体乳酸脱氢酶有5条同工酶谱带,由LdhA、LdhB两个基因编码。该研究可为扁圆吻鲴种质评估及种质标准制定提供理论参考。
参考文献
[1]
王昕阳.细鳞斜颌鲴人工繁育及养殖技术[J].黑龙江水产,2017(6):17-19.
[2] 杨品红,徐黎明,王志陶,等.细鳞斜颌鲴不同组织中LDH同工酶的比较研究[J].激光生物学报,2009,18(4):509-515.
[3] 何舜平,刘焕章,陈宜瑜,等.基于细胞色素b基因序列的鲤科鱼类系统发育研究(鱼纲:鲤形目)[J].中国科学(C辑:生命科学),2004,34(1):96-104.
[4] 李行先,杨仲景,冯晓宇,等.圆吻鲴人工繁殖和苗种培育[J].杭州农业科技,2004(1):15-17.
[5] 何美峰,袁定清,崔利峰,等.梯级电站胁迫下旧县河下游鱼类群落多样性分析[J].福建水产,2014,36(3):198-204.
[6] 林义浩.快速获得大量鱼类肾细胞中期分裂相的PHA体内注射法[J].水产学报,1982,6(3):201-208.
[7] 郭丰,王军,苏永全,等.云纹石斑鱼染色体核型研究[J].海洋科学,2006,30(8):1-3.
[8] 贺刚,方春林,王伟萍,等.大口鲇(♀)×怀头鲇(♂)杂种F1的形态学及遗传学研究[J].湖北农业科学,2011,50(23):4916-4919.
[9] 孟彦,张燕,许映芳,等.月鳢和乌鳢同工酶的比较研究[J].安徽农业科学,2009,37(27):12951-12952,12964.
[10] 余来宁,夏小平,杨东,等.草鱼♀×鳡♂杂交F1代同工酶和蛋白质的电泳分析[J].安徽农业科学,2014,42(30):10573-10575,10628.
[11] 乔德亮,李思发.细鳞斜颌鲴三个群体形态差异[J].生态学杂志,2010,29(12):2425-2430.
[12]WIMBERGER P H.Plasticity of fish body shape.The effects of diet,development,family and age in two species of Geophagus(Pisces: Cichlidae)[J].Biological journal of the linnean society,1992,45(3):197-218.
[13] 张林,周剑光,张涛,等.长江水系细鳞斜颌鲴形态特征及生化遗传特性分析[J].中国渔业质量与标准,2018,8(2):29-35.
[14] 李康,李渝成,桂建芳,等.中国鲤科鱼类染色体组型的研究——Ⅸ.鳊亚科9种鱼和鲴亞科1种鱼的染色体组型[J].水生生物学报,1986,10(2):189-193.
[15] 秦改晓,张潇,齐子鑫,等.人工养殖黄河鲤形态特征、染色体核型及肌肉乳酸脱氢酶的电泳分析[J].南方农业学报,2019,50(8):1844-1850.
[16] 洪云汉,周密,周暾.中国鲤科鱼类染色体组型的研究——(Ⅲ).鳑鲏亚科七种鱼的染色体的比较分析[J].武汉大学学报(自然科学版),1983,29(2):96-102,116.
[17] 马纲.中国淡水鱼类染色体形态及数目变异的研究进展[J].甘肃科学学报,1996,8(3):77-80.
[18] 章海鑫,徐先栋,张燕萍,等.黄尾鲴染色体核型分析[J].江西水产科技,2018(5):20-22.
[19] MARKERT C L,SHAKLEE J B,WHITT G S.Evolution of a gene.Multiple genes for LDH isozymes provide a model of the evolution of gene structure,function and regulation[J].Science,1975,189(4197):102-114.
[20] 吴力钊,王祖熊.白鲢个体发育过程中同工酶基因的表达与调控研究[J].水生生物学报,1997,21(1):49-58.
[21]
SHAKLEE J B,KEPES K L,WHITT G S.Specialized lactate dehydrogenase isozymes:The molecular and genetic basis for the unique eye and liver LDHS of teleost fishes[J].Journal of experimental zoology,1973,185(2):217-240.
[22] TURAN C.Stock identification of Mediterranean horse mackerel (Trachurus mediterraneus) using morphometric and meristic characters[J].ICES Journal of Marine Science,2004,61(5):774-781.
