抗原设计新技术及其在疫苗中的应用和发展
2021-04-23李兴航综述杨晓明审校
李兴航 综述,杨晓明 审校
1.国家联合疫苗工程技术研究中心,湖北武汉430207;2.武汉生物制品研究所有限责任公司,湖北武汉430207;3.中国生物技术股份有限责任公司,北京100029
自1798 年首次发现接种牛痘可以用来防治天花开始,开启了人类研发和应用疫苗来预防疾病的时代。过去200 多年来,疫苗的应用降低了多种传染性疾病的发病率和死亡率。据报道,疫苗每年可减少全世界约250 万人死亡[1],大大减低了由传染性疾病所带来的公共卫生负担。
疫苗发展初期,是由詹纳等人开创的研究疫苗的方法,目前被认为是传统的疫苗学(也被称作“疫苗学1.0”)[2]。他们当时并不知道病原体的具体来源及疫苗发挥作用的机制,而是通过观察发现使用奶牛的脓包(牛痘)可以预防天花,并在人群中进行试验,结果获得了极好的效果。“疫苗”从此诞生,这是人类研究疫苗历史上的一座里程碑。接下来,巴斯德传承了詹纳对疫苗接种的概念,又进行了创新,于1885 年成功研制出第一款人用狂犬病疫苗。后来,人们总结出巴斯德研究疫苗的3 个定理:分离、灭活、注射,这开始引导研究者更加合理地设计疫苗[2]。第一代疫苗基本遵从了巴斯德的疫苗设计策略,包括天花疫苗、卡介苗(BCG)、鼠疫疫苗、百日咳疫苗等。“疫苗学 2.0”兴起于 20 世纪 90 年代[3],这个转折点是b 型流感嗜血杆菌(haemophilus influenzae type B,Hib)和肺炎链球菌疫苗[4]的问世。第二代疫苗开始对病原体的致病机制有了一定的了解,并结合新的方法和技术(如遗传学、蛋白质工程、重组DNA技术、多糖化学技术、合成生物学[5]等相关知识)对细菌和病毒成分进行分离和纯化。亚单位疫苗和多糖蛋白结合疫苗即为“疫苗学2.0”时期出现的成果。目前的破伤风、流行性感冒、白喉、炭疽、肺炎、乙肝等疫苗均是第二代疫苗的代表。
传统疫苗设计方法研制的疫苗对于乙肝、狂犬病、破伤风等疾病的控制起了很大作用,大大降低了这些疾病在人群中的感染率。但目前传统研究方法在某些方面还有待进一步突破,如①无法应用于体外不可培养的微生物[如丙型肝炎病毒(hepatitis C virus,HCV)、人乳头瘤病毒(human papillomavirus,HPV)][6];②无法为易发生突变的病原体[如甲型流感病毒(influenza A virus,IAV)、人类免疫缺陷病毒(human immunodeficiency virus,HIV)]提供广泛的交叉保护[7];③对于某些疾病(如登革热、疟疾)尚不能提供很好的解决方案[8];④无法用于机会致病菌(如金黄色葡萄球菌)[9]。
目前,基因组学、免疫组学、蛋白质组学、结构疫苗学等新兴科学相继出现并相互融合,共同推动了对传统疫苗设计思路的创新[6]。这些新兴学科的出现开创了疫苗设计的一个新时代—“疫苗学3.0”时代,为针对如HIV、IAV、呼吸道合胞病毒(respiratory syncytial virus,RSV)、登革热病毒(Dengue virus,DENV)等病原体疫苗的设计和开发开辟了新的思路和对策。本文就抗原设计新技术及其在疫苗中的应用和发展作一综述。
1 结构疫苗学设计疫苗的理论基础及策略
1.1 结构疫苗学的理论基础 由于抗原特异性所诱导的体液免疫与很多感染性疾病的保护有关[10],一直以来,人类研发疫苗的目标之一即是要诱导出对机体具有保护作用的特异性抗体,且能在体内维持较长时间。在疫苗的抗原设计中,要想该抗原在人体内诱导出针对某病原体的特异性抗体,即需对该病原体特定分子的精确结构进行分析。于是,研究者开始将研究方向转向抗原的结构与疫苗的关系上。
但一些病原体可通过表面残基多样化(如HIV)使之前研发的疫苗失去保护作用;有些病原体的保护性抗原处于一种亚稳态的构象(如RSV),即其结构在感染过程中会发生改变,逃避由疫苗诱导的免疫应答;此外,还有些病毒的抗原(如DENV、寨卡病毒)经免疫应答产生的抗体甚至会加重疾病,出现抗体介导的增强感染(antibody-dependent enhancement,ADE)现象。因此,迄今尚未研究出效果良好的针对RSV、HIV、DENV 等病原体的疫苗。目前研究者开始考虑是否可通过在结构上对抗原进行修饰和改造,设计出有一定理化特性的抗原表位,诱导出针对某病原体的特异性抗体。结构疫苗学(structuralvaccinology,SV)[11]应运而生,蛋白质工程技术开始被应用于疫苗抗原的设计中。
现有的结构生物学方法已能够确定全病毒、病毒蛋白以及抗原-抗体复合物的结构[12]。对病原体的三维(3D)结构分析能够分析该病原体上各抗原三级结构的信息和位置。这些结构信息可被用来解决目前阻碍疫苗开发的一些问题。运用结构疫苗学设计候选疫苗抗原的主要步骤如下[13]:①确定抗原或抗原-抗体复合物的结构;②通过分子生物学手段重新构建抗原或表位;③将重组抗原或表位应用于疫苗平台上;④测试候选疫苗在动物模型上的安全性和有效性(图1)。简而言之,结构疫苗学将基因工程与高分辨率的结构分析技术相结合,使得病原体的保护性抗原成分能够被选择性地构建,并应用于疫苗中。

图1 结构疫苗学中基本的方法和工具Fig.1 Basic methods and tools in structural vaccinology
1.2 结构疫苗学中抗原设计的策略及运用
1.2.1 以表位为中心的疫苗设计策略 病原体上不同的抗原表位对于机体有不同的免疫原性,即不同表位的免疫优势级别也不同,其中一些抗原表位可优先激活生发中心的B 细胞[14-15]。以表位为中心的策略重点在于将免疫原中与保护性抗原成分不相关的抗原表位去除,使得B 细胞能聚焦到识别和提呈可诱导针对病原体产生保护性中和抗体的抗原表位。如流感病毒上至少有10 种病毒蛋白,但只有血凝素(hemagglutinin,HA)对疫苗的设计至关重要,目前流感疫苗所诱导的中和抗体均是针对HA 蛋白的顶部区域。但运用蛋白质工程技术有3 种方法对抗原进行设计[11]:①表位移植;②结构域最小化;③表面重构。
表位移植[11,16]是将抗原天然构象中能诱导机体产生中和抗体的表位移植至其他异源蛋白上的过程。该方法早期应用于RSV 疫苗的研发中。CORREIA 等[17]开发了一种名为 Fold From Loops(FFL)的计算方法,并设计了一种包含RSVF 蛋白抗原位点Ⅱ的蛋白质支架,使其呈现与RSV 晶体结构中观察到的几乎相同的构象,最终的结果未能在小鼠中诱导RSV 中和活性,但能在猕猴中诱导大量的中和抗体。虽然未取得理想的结果,但这些尝试仍为以表位为中心的疫苗设计提供了宝贵的经验。另一个应用的实例是针对B 群脑膜炎球菌(meningococcus,MenB)疫苗的抗原设计。HOLLINGSHEAD 等[18]基于结构设计了一种嵌合抗原(Chimeric antigens,ChAs),用 H 因子结合蛋白(factor H binding protein,fHbp)作为蛋白质支架,在支架上嵌合了来自整合膜蛋白PorA中的VR2 表位。PorA 具有较高的免疫原性,其上的VR2 是一个可在人和动物模型中诱导产生杀菌性抗体的表位[19-20]。最后将这种嵌合抗原免疫小鼠模型,在小鼠体内成功诱导出了针对fHbp 和PorA 蛋白的抗体。
结构域最小化[11]是将含多个结构域的蛋白质改造为仅携带中和抗原表位的单一结构域的过程。如运用该原理设计的通用流感疫苗,是将流感病毒表面HA 头部下面较为保守的的茎部暴露出来,使体液免疫应答能集中于此[21-22]。YASSINE 等[23]首先对流感病毒的HA 在3D 结构上进行分析,借助计算机技术对HA 蛋白通过6 次连续改造,构建出mini-HA 茎(mini-HA stem)抗原,使得茎部保守区占整个HA 表面面积的比例由最初的37%显著提升为94%。在雪貂和小鼠模型上进行免疫,均可检测到针对流感病毒的广谱中和抗体。
表面重构[11]是将与能诱导保护性免疫应答不相关的表位改变为非优势表位的过程。如在HIV疫苗开发遇到的一个问题:HIV 包膜上的许多表位诱导非中和抗体,这些非中和抗体会阻碍中和抗体发挥作用[24]。WU 等[25]使用 HIV-1 完整包膜结构的信息开发了一种表面被重新设计的糖蛋白,这些糖蛋白诱导的免疫应答可特异性地针对初始CD4受体结合位点保守位点的抗原。NACHBAGAUER等[26]研究了一种嵌合的 HAs(cHA),将一种不在人体群中循环的流感病毒的 HA 头部(H5、H8、H9)与流感流行株的HA 茎部(H1)进行组合(图2),可选择性地增强诱导对茎部结构的抗体。并在雪貂动物模型中成功诱导了广泛的交叉反应性抗体。此外还发现,用这种嵌合抗原初次免疫,再用裂解流感疫苗进行加强免疫,获得了比连续接种两次裂解疫苗更优异的针对H1N1 流感病毒的保护效果。
以表位为中心的设计策略中,表位移植、结构域最小化和表面重构3 种方法可根据不同病原体的抗原仅使用一种策略,也可联合多个策略使用以提高抗原的免疫原性。

图2 嵌合HA 抗原结构图Fig.2 Structure of chimeric HA antigen
1.2.2 稳定抗原的天然构象 病原体感染人体后,其中的某些蛋白质可能会发生构象的改变,而构象稳定技术可将这类表位固定为可诱导保护性抗体的状态。如RSV 的F 蛋白,在感染过程中从亚稳定性的融合前构象重新排列成高度稳定的融合后构象(图3[13])。而融合前 F 蛋白表位被视作 RSV 疫苗及药物研究的重要靶点,因为该表位能够诱导人体产生针对RSV 的保护性中和抗体来预防RSV 感染[27-28]。ROSSEY 等[29]、GRAHAM 等[30]和 MCLELLAN 等[31]通过构象稳定技术将融合前F 蛋白固定。其大致步骤是将T4-噬菌体纤蛋白三聚化结构域(“foldon”)添加至 RSV F 膜外结构域[32]的 C-末端,并结合融合前特异性D25 抗体。并与其他方法同时运用(包括引入半胱氨酸对、添加二硫键、空腔填充)来稳定其结构。用该候选抗原免疫动物后,在动物体内检测到了较高的保护性抗体滴度[33]。
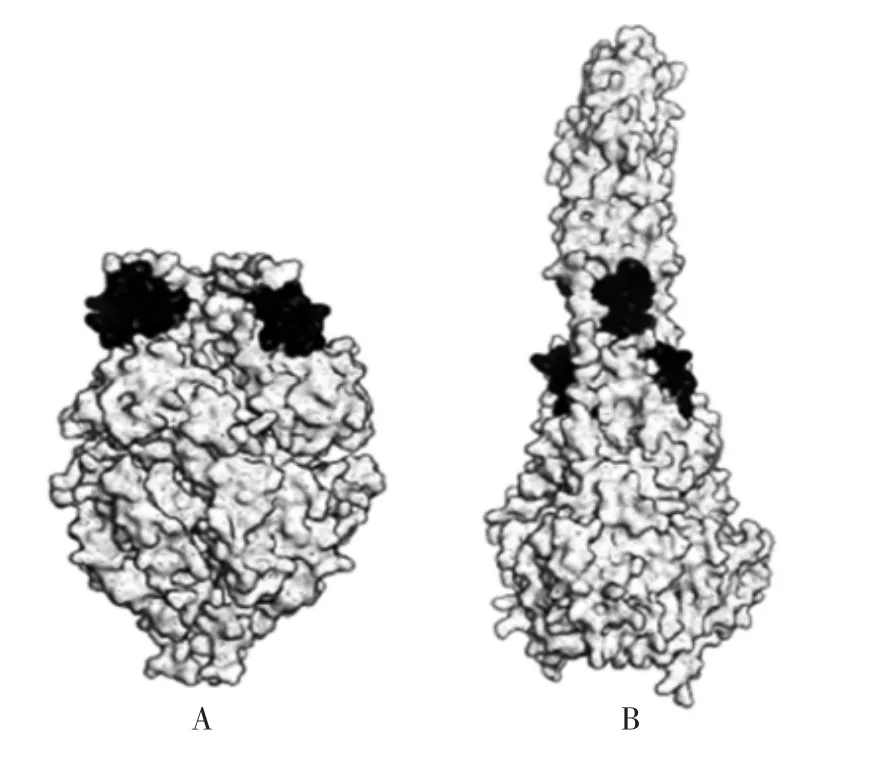
图3 RSV 融合前(A)后(B)F 蛋白构象示意图Fig.3 Confirmation of F protein of RSV before(A)and after(B)fusion
1.2.3 构建模拟病原体的B 细胞表位 一些病原体逃避宿主免疫的机制是序列多样化,如HIV 的包膜蛋白、流感病毒的HA 蛋白。尽管这些位点在疫苗的设计中被非常重视,但由于其多样性,很难研发出单一的疫苗来预防这种疾病。为此,可通过设计模拟病原体的B 细胞表位,并在该表位中引入某抗原的共有序列以诱导合成一种广谱的中和抗体。以A 型流感病毒为例,目前上市的疫苗由于交叉保护水平低,必须每年根据预测当年流行株进行更新[34]。因此有个假设,在多个亚型的流感病毒HA 蛋白的基因序列中,发掘出一段相同或相似的序列片段,将这部分序列对应的蛋白作为疫苗候选抗原,就能研究一种通用流感疫苗。LADDY 等[35]在 H5N1 病毒的研究中报道了一种获取流感病毒表面HA 蛋白的共有序列的方法,并在小鼠、雪貂、非人类的灵长类动物3 种动物模型中得到了针对H5N1 流感亚型不同病毒株的中和抗体。之后,YAN 等[36]也报道了一种微共有序列(micro-consensus)应用于疫苗领域的案例,他们通过对近25 年来流行的H1 流感亚型的HA 基因序列进行分析鉴定,得出4 个主要的基因簇,并基于此设计出包含这4 个主要基因簇的微共有序列的DNA 疫苗。在小鼠、豚鼠和非人类的灵长类动物模型中的研究发现,该DNA 疫苗能诱导出对近10 年流行的H1 亚型流感病毒具有保护性的广谱中和抗体。
1.2.4 多价纳米颗粒 与单体抗原不同,多价纳米颗粒可快速穿梭至滤泡树突状细胞(follicle dendritic cell,FDC)网络,然后触发补体的甘露糖结合凝集素(mannose-binding lectin,MBL)途径和免疫原聚糖依赖性方式在生发中心集中诱导免疫应答。如针对HIV-1,TOKATLIAN 等[37]检测了两种 HIV 包膜抗原作为可溶性“单聚体”或蛋白质纳米颗粒在动物模型中的免疫效果,分别为gp120 的种系靶向工程外部结构域(eOD-GT8,简称 eOD)和gp140 包膜三聚体(MD39)。与可溶性免疫原相比,纳米颗粒形式的eOD 和MD39 免疫小鼠能诱导更高的IgG 滴度(高达90 倍)。应用纳米颗粒的其他候选疫苗抗原在动物模型中的研究结果也表明,与单体抗原相比,经纳米颗粒修饰的免疫原能更有效地激活低亲和力的前体 B 细胞[38-40],增强的滤泡辅助 T 细胞(follicular T-helper-cell,Tfh)诱导与生发中心(germinalcenter,GC)反应,并增强中和抗体的诱导[41]。总之,使用纳米颗粒作为骨架,并以此为基础对抗原进行装载和修饰已被证明是一种抗原设计的可行手段。
1.3 结构疫苗学的局限性 目前来看,结构疫苗学具有巨大的潜力,但也有一些局限性。首先,目前研究者尚未完全掌握由病毒诱导的机体免疫应答过程中的结构和免疫学知识,该技术可能会因此受到影响。如在DENV 包膜蛋白的DⅢ结构域上有一个结构保守、对温度敏感、隐蔽的表位,但该表位在DENV 的包膜蛋白 E 抗原上很难被发现[42]。LI 等[42]用3E31 抗体阻止了E 抗原介导的膜融合,从而间接证明该抗体识别了DENV 表面E 抗原上尚未了解的中间构象。
其次是X 射线晶体学只能分析抗原-抗体复合物的静态结构[43]。但当抗原-抗体相互作用时,它们之间是相互适应的动态过程。人们无法通过静态结构很好地推测其动态过程中的细节。因此,通过该技术观察到的特定表位结构不一定能对应于所设计出的产生抗体的表位,这些表位不一定是应用在疫苗中的最佳表位。VAN 等[44]发现,病毒的三聚体Env 具有巨大的可塑性和结构灵活性,因此很难确定Env 的哪些结构域是最有可能诱导保护性抗体的最佳候选疫苗免疫原。
最后,结构疫苗学的应用目前局限于重组蛋白疫苗、肽疫苗和病毒样颗粒(virus-like particles,VLPs)疫苗的设计,这些疫苗平台的免疫原性有待提升。因此,将结构疫苗学与其他技术结合运用也是未来可以考虑的发展方向。目前的结构疫苗学非常重视B细胞介导的体液免疫,但对T 细胞介导的细胞免疫方面尚未深入研究,未来的发展可考虑对疫苗诱导的细胞免疫进行研究。
2 疫苗组学在候选抗原发现和筛选中的应用
目前已进入一个大数据的时代,支持高通量技术的组学学科(如基因组学和后基因组学[45-46],包括转录组学、蛋白质组学、代谢组学、细胞组学、免疫组学、信息组学等),能生成和处理大量的数据和信息。于是,疫苗组学的概念被提出,这为疫苗的候选抗原发现及筛选开辟了另一条途径。
反向疫苗学(reverse vaccinology,RV)是其的一个代表,而且已被证明是非常有效的手段。MenB 疫苗的抗原设计就是运用反向疫苗学策略对疫苗候选抗原进行选择。该策略首先通过对病原体进行全基因测序,并对基因组所编码的所有蛋白质进行克隆,再对每种蛋白质进行鉴定和筛选。由于该过程涉及动物免疫,待测抗原的数量越多,整个过程将会非常繁杂。如在MenB 的疫苗抗原筛选中,PIZZA 等[47]首先从MenB 基因组编码的总计2 000 余种蛋白质中筛选出570 种表面蛋白(因为他们认为只有表面抗原才可能成为保护性抗原),再对这570 种蛋白质逐个进行筛选鉴定,最后确定出3 种蛋白作为疫苗候选抗原。尽管只对570 种蛋白进行了筛选,但整个过程仍非常费时费力。
为了避免这些费时的工作,需要新的生物信息学工具和方法来更好地整合来自组学实验的大量数据,使得从单组学向多组学的转变成为可能。如计算疫苗学[48]和免疫信息学[49]利用算法,可让人们仅关注整个病原体中部分筛选过的抗原,缩减整个筛选中耗时和劳动密集的步骤,从而降低成本。
DOROSTI 等[50]运用疫苗组学策略研究了一种肺炎链球菌亚单位疫苗,包含了一些保守毒力蛋白的混合物,打破了常规用荚膜多糖作为抗原的传统疫苗。该研究分为3 步:首先,检索了肺炎链球菌PspA、CbpA、PhtD 和 PiuA 的氨基酸序列,这些表位来源于不同数据库的筛选(如IED8、PROPRED、RANKPED 和 MHCPRED)。PspA 和 PcbA 用作 CTL 表位刺激剂,PhtD 和PiuA 用作辅助表位,并使用PorB蛋白作为TLR2 激动剂来增加疫苗的免疫原性。然后通过适当的氨基酸将辅助抗原、靶向抗原和TLR激动剂融合。并对其理化、结构和免疫学特性进行评估。最后,用 Jcat(http:/ / www.jcat.de)对输出DNA 序列进行优化和反向翻译,并在大肠埃希菌中克隆和表达。还可通过大数据信息对特定病原体的基因进行分析,以预测其具有跨膜结构域的蛋白质、前导肽、膜锚定结构域等。MASIGNANI 等[51]就是基于此在脑膜炎奈瑟球菌表面发现了一种新的脂蛋白GNA1870,其可诱导机体产生保护性的杀菌抗体。
为了进一步缩减人力物力成本,使待测候选抗原的范围再次缩小,ALTINDIS 等[52]提出一种“保护组学分析”的策略。他们不再关注蛋白质定位,而是将其计算分析集中在蛋白质的生物学作用和功能上。他们认为,那些能诱导机体产生保护性抗体的蛋白质抗原在结构或功能上具有某种相同的特性。这种策略可大大减少实验中候选抗原的数量,大概仅为某种病原体全部蛋白数量的3%。不仅如此,由于该策略基于的是抗原的结构与功能进行筛选,而并不局限抗原所在的位置,保护组学分析能发现传统研究方法关注不到的一些胞内抗原。
3 展 望
传统研究疫苗的方法是对病原体进行灭活或减毒,后来免疫学和分子生物学等学科的发展有力地扩充了疫苗的研究策略。目前市兽疫苗均是基于这些策略研究的,有效控制了人类多种传染病的传播,具有十分重要的社会效益和经济价值。但针对艾滋病、登革热、疟疾等对人类生命安全有重大威胁传染病的病原体,目前尚未研发出有效的疫苗。此外,不断有新的传染病出现,对疫苗的研发也是一种挑战,如之前发生在中国的SARS 以及最近在非洲暴发的埃博拉疫情,造成了大量生命和财产损失。因此,在肯定传统疫苗作用的同时,新型疫苗的研发也迫在眉睫。
目前,结构疫苗学和疫苗组学等疫苗设计新策略的发展,对于传统疫苗设计策略难以解决的问题已有了新的解决思路。针对的病原体也不局限于文中提到的,如最近针对疟疾[53]、霍乱弧菌[54]、马尔堡病毒[55]的疫苗设计均用到了上述提到的新的抗原设计策略。
由于目前疫苗开发的大都是由抗体以及其靶标的结构所驱动的,但不同病原体使用不同的策略来逃避保护性的免疫应答,因此,疫苗设计策略必须根据对每种疾病中保护性抗体如何发挥其功能,以及特定抗原如何诱导免疫进行调整[56]。并可能需要结合多种策略来设计针对某一种病原体的疫苗,而不是单独的依赖某一项技术或策略。另外,在应用新技术对下一代疫苗进行设计的同时,不仅要考虑其抗原的设计,还要考虑抗原的投递途径[57-58],疫苗在运输中抗原的稳定性[13]等问题,在研发出更稳定、高效的新一代疫苗的同时,能够让更多的人接种到疫苗。
