陆地棉纤维起始期优势表达基因GhCRPK1 的克隆及功能研究
2021-04-16王雪慧陈丽锦赵若林程海亮张友平王巧连吕丽敏宋国立左东云
王雪慧,陈丽锦,赵若林,程海亮,张友平,王巧连,吕丽敏,宋国立,左东云
(中国农业科学院棉花研究所/ 棉花生物学国家重点实验室,河南 安阳 455000)
关键字:类受体激酶;GhCRPK1;纤维发育;表皮毛;信号传导
植物类受体激酶(Receptor like kinase,RLK)是一类跨膜受体蛋白, 通常具有胞外结构域、跨膜结构域及胞内的激酶结构域。 胞外结构域可以感知细胞外或环境中的信号分子,跨膜结构域将受体激酶镶嵌到细胞膜上,胞内激酶结构域具有丝氨酸/苏氨酸磷酸化位点, 能将底物磷酸化传递信号。 植物中RLKs 的胞内激酶结构域具有较高的同源性, 但是胞外结构域具有较大的差异。根据胞外结构域的序列特点,可以将RLKs 分为亮氨酸重复序列(Leucine rich-repeat,LRR)受体激酶、凝集素(Lectin)受体蛋白、表皮生长因子(Epidermal growth factor,EGF)受体蛋白、细胞壁偶联受体激酶 (Wall-associated kinase,WAK)及受体类细胞质激酶(Receptor-like cytoplasmic kinase,RLCK)等[1]。 RLCK 没有胞外结构域只含有胞内激酶结构域,一般通过与其它RLKs 相互作用进行信号传递。
研究表明,受体激酶RLKs 在植物的生长发育及防御反应中发挥着重要的作用。 FLS2(Flagellin-sensitive 2) 能够识别细菌的鞭毛蛋白N 末端的22 个氨基酸,激发拟南芥的免疫反应,提高拟南芥对细菌的抗性[2-4]。延伸因子EF-Tu 受体(EFTu receptor,EFR) 能识别细菌的延伸因子EF-Tu(Elongation factor thermo unstable),增强拟南芥对农杆菌的抗性[5]。 拟南芥BRI1(Brassinosteroid insensitive 1) 能够识别油菜素内酯(Brassinos teroids,BRs),然后激活BR 信号通路[6-7]。 CLAVATA1 是1 个在分生组织中表达的LRR 类受体激酶, 识别CLAVATA3 短肽信号分子后, 抑制WUSCHEL(WUS)基因表达,调控分生组织的形成[8-9]。 FER(FERONIA)是1 个长春花类受体激酶(Catharanthus roseusRLK1,CrRLK1)蛋白,其对应的信号分子是RALF (Rapid alkalinization factor),调控植物根的发育及花粉的生长[10-13]。 此外,ANX1(ANXUR1)、ANX2(ANXUR1)、BUPS1(Buddha’s Paper Seal l)、BUPS2(Buddha’s Paper Seal 2)及LRX(Leucine rich-repeat extensin) 能够识别RALF 家族的其他成员进而调控花粉管的生长及花粉管与卵细胞的识别[14-18]。RLKs 参与植物生长发育及抗逆反应的各方面,是胞外信号与胞内应答反应的桥梁,是植物响应各种环境信号的重要组成部分。
棉花是最重要的天然纤维的来源,在我国国民经济中占有举足轻重的地位。陆地棉(Gosspium hirsutum)是主要的棉花栽培品种,产量高、适应性广,占棉花总产的95%以上。 尽管在拟南芥和水稻等植物中鉴定了部分RLKs 的生物学功能,但是RLKs 在棉花中的功能研究较少,特别是参与棉花纤维发育的机制知之甚少。 本研究基于陆地棉全基因组关联分析的最新数据[19],并利用荧光定量聚合酶链式反应 (Quantitative real-time polymerase chain reaction,qRT-PCR)的方法筛选到在棉花纤维发育起始期优势表达的基因GhCRPK1,推测该基因可能参与了棉花纤维发育的起始, 在拟南芥中验证了该基因的分子功能,并利用酵母双杂交的方法筛选了与这个基因相互作用的蛋白,为揭示RLKs 调控棉花纤维发育的分子机制提供理论基础。
1 材料与方法
1.1试验材料
试验所用的棉花材料陆地棉遗传标准系TM-1、拟南芥哥伦比亚野生型(Col-0)、本氏烟草均由本课题组保存,拟南芥突变体cpk1-1(SALK_004253C)和cpk1-2(SALK_054771)由中国农业大学杨淑华教授馈赠。 大肠杆菌DH5ɑ 和农杆菌GV3101 购自上海唯地生物技术有限公司;PCR扩增采用南京诺唯赞生物科技有限公司的高保真酶2×PhantaMax Master Mix;RNA 提取使用天根生化有限公司的多糖多酚植物总RNA 提取试剂盒;cDNA 反转录采用宝生物的PrimeScriptTM1st Strand cDNA Synthesis Kit 反转录试剂盒;qRT-PCR 分析使用Genstar 公司的2×RealStar Green Fast Mixture(with ROX II);同源重组采用南京诺唯赞生物科技有限公司的ClonExpressUltra One Step Cloning Kit 试剂盒。
1.2 试验方法
1.2.1取样、RNA 提取、逆转录和荧光定量分析。陆地棉标准系TM-1 种植在中国农业科学院棉花研究所老所部农场(河南省安阳县白璧镇),在开花当天挂牌标记,分别采集开花前5 d(-5 days post anthesis,-5 DPA),开花前3 d(-3 DPA)、开花前1 d(-1 DPA)、开花当天(0 DPA)、开花后1 d(1 DPA)、开花后3 d(3 DPA)、开花后5 d(5 DPA)、开花后7 d(7 DPA)和开花后10 d(10 DPA) 的棉铃, 其中10 DPA 的棉铃拨取纤维、 其他时期取的是胚珠和纤维的混合物。 在ABI QuantStudio 3 实时定量PCR 仪上进行qRT-PCR 分析,所有操作均严格按照试剂盒使用说明进行。 内参基因为Histone 3,所有基因的引物见附表1。 qRT-PCR 反应体积为20 μL,扩增程序:95 ℃预变性2 min;95 ℃变性15 s、60 ℃退火15 s、72 ℃延伸20 s, 采集荧光,40 个循环后绘制熔解曲线。
1.2.2GhCRPK1 基因克隆。 根据GhCRPK1基因的序列信息, 设计引物GhCRPK1PF1/GhCRPK1PR1(附表1),以TM-1 的cDNA 为模板扩增GhCRPK1基因,将PCR 产物切胶回收克隆到pCE-Zero 载体上, 转化大肠杆菌DH5ɑ 感受态,挑选阳性克隆,送往上海生工生物技术有限公司测序。
1.2.3生物信息学分析。 从Cotton FGD 网站(https://cottonfgd.org/)获取Gh_A10G1849 的基因序列和蛋白质序列, 并以Gh_A10G1849 的蛋白序列作为查询序列搜索并下载海岛棉、亚洲棉和雷蒙德氏棉中的同源蛋白序列;同时在植物基因组网站(http://ensemblgenomes.org/)通过BLASTP搜索并下载番茄、拟南芥、水稻、小麦、玉米等物种中的同源蛋白序列,在各物种中选择同源性最高的1~2 个蛋白进行蛋白保守性分析和进化树分析。 使用ClustalX[20]软件进行蛋白多序列比对分析,用MEGAX[21]软件构建系统进化树。
1.2.4亚细胞定位分析。 在GhCRPK1编码序列前端和末端分别添加SalI 和BamHI 酶切位点,在GFP序列前端和末端分别添加BamHI 和EcoRI 酶切位点,扩增引物为GhCRPK1PF2/GhCRPK1PR2 (附表1)。 通过同源重组构建pCaMV35S∷GhCRPK1-GFP 载体, 测序结果比对正确后转化到农杆菌菌株GV3101,注射生长1 月左右的烟草幼嫩叶片。黑暗条件下室温处理2 d 后,在激光共聚焦显微镜(Leica TCS SP8)下观察荧光。
1.2.5拟南芥过表达及表皮毛观察。 PCR 扩增GhCRPK1基因的编码区, 扩增引物为GhCRPK1PF3/GhCRPK1PR3(附表1),通过同源重组构建GhCRPK1过表达载体pCaMV35S::GhCRPK1-His。 载体测序结果比对正确后转化到农杆菌菌株GV3101,利用浸花法转化拟南芥[22],并在含有50 ng·mL-1卡那霉素的MS 培养基中筛选阳性植株。 选取4 周大小的拟南芥植株莲座叶, 在无水乙醇中脱色2 h 后, 在体式显微镜(Olympus SZX10) 下观察拟南芥表皮的形态,按照参考文献[19]中的方法统计拟南芥表皮毛的长度和密度。
1.2.6酵母双杂交文库筛选及互作蛋白点对点验证。 将GhCRPK1基因的编码序列插入到pGBKT7 载体中,筛选陆地棉纤维发育起始时期的酵母表达文库。 将筛选到的阳性克隆测序后从陆地棉中克隆其编码序列构建到pGADT7 载体中,与pGBKT7-GhCRPK1 进行点对点的互作验证。 将筛选得到的Gh_D08G0057 编码序列连接到pGADT7 载体中,分别将pGBKT7-GhCRPK1和pGADT7-T 质粒(对照)及pGBKT7-GhCRPK1和pGADT7-Gh_D08G0057 质粒共转化酵母,分别在二缺培养基 (无亮氨酸和色氨酸,即SD/-Leu/-Trp,SD-DDO)和添加α- 半乳糖苷酶显色底物(X-α-gal)、抗生素金担子素A(Aureobasidin A,AbA)的四缺培养基(无亮氨酸、色氨酸、组氨酸和腺嘌呤, 即SD/-Leu/-Trp/-His/-Ade,SD-QDO)中生长。所有操作均严格按照Clontech公司的MatchmakerGold Yeast Two-Hybrid System 说明进行。
2 结果与分析
2.1 与棉花纤维发育相关的受体激酶基因的筛选
为了筛选陆地棉中与纤维发育相关的受体激酶,利用陆地棉全基因组关联分析数据,筛选到34 个与棉花纤维长度、 强度和衣分相关的受体激酶候选基因,这些基因可能在棉花纤维发育过程中发挥着重要的作用。 利用qRT-PCR 分析这34 个基因在棉花纤维发育不同时期的表达量发现, 其中18 个基因在棉花纤维发育过程中的表达具有一定的规律(图1),而其他基因在棉花纤维发育过程中的表达没有明显规律。 这18 个基因在纤维发育过程中具有不同的表达模式:部分基因如Gh_D08G2087、Gh_A13G2073、Gh_A08G2425、Gh_D02G0129、Gh_A06G1068、Gh_A08G0809、Gh_A09G0625、Gh_A10G1831、Gh_A08G2273、Gh_A01G0896、Gh_D13G2124和Gh_D08G00480在开花后5 d、7 d 或10 d 优势表达(图1A);部分基因Gh_A10G0312、Gh_A09G0186和Gh_A08G0234 的表达量在开花前较高, 在开花后降低(图1B);部分基因在棉花纤维发育的起始时期表达水平较高, 如Gh_A10G1849在开花当天及开花后1 d 和3 d 表达水平高,Gh_D05G3666在开花前1 d 表达水平最高,Gh_A09G0177在开花后1 d 表达水平最高(图1C)。Gh_A10G1849、Gh_A09G0177和Gh_D05G3666三个基因在纤维发育起始时期高水平表达,表明其可能参与了棉花纤维发育起始, 特别是Gh_A10G1849在开花当天到开花后3 d 持续高水平表达, 与棉花纤维发育起始的时期一致,推测其在棉花纤维发育起始过程中发挥着重要的作用。
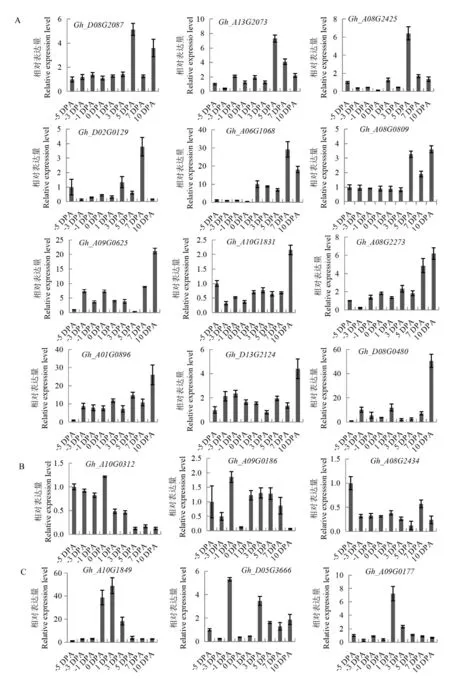
图1 18 个受体激酶基因在棉花纤维发育不同时期的表达模式分析Fig. 1 Expression patterns of 18 receptor kinase genes at different stages of cotton in fiber development
2.2 Gh_A10G1849 基因的克隆及生物信息学分析
在CottonFGD 数据库中搜索Gh_A10G1849基因,发现该基因的全长为4 594 bp,包含有1 个1 047 bp 的开放阅读框, 编码348 个氨基酸,预测蛋白的分子量为39.11 KDa, 等电点为7.34。在TM-1 中克隆该基因完整的编码序列, 经过测序比对, 与数据库中Gh_A10G1849基因的序列完全一致。 对该基因进行功能注释发现其与拟南芥冷响应蛋白激酶1 (Cold-responsive protein kinase 1,CRPK1) 编码基因At1g16670同源性较高,因此将该基因命名为GhCRPK1。 CRPK1 属于RLK 家族中的RLCK 亚家族, 无胞外结构域和跨膜结构域。 同源蛋白序列分析发现,CRPK1在二倍体棉花雷蒙德氏棉和亚洲棉、四倍体棉花陆地棉和海岛棉中均存在,并且广泛存在于拟南芥、番茄和可可等双子叶植物及水稻、玉米和小麦等单子叶植物中, 表明CRPK1 可能在植物中发挥重要的作用。
比较陆地棉中的GhCRPK1 与海岛棉(G.barbadense)、亚洲棉(G. arboreum)、雷蒙德氏棉(G.raimondii)、 可可(Theobroma cacao)、 番茄(Solanum lycopersicum)、 拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、玉米(Zea mays)和小麦(Triticum aestivum)中的CRPK1 的蛋白序列,尽管存在一定的差异,但是它们发挥功能的激酶结构域都是高度保守的(图2)。 进一步分析这些物种中的CRPK1 蛋白的进化关系发现 (图3),CRPK1 蛋白在进化上也具有保守性, 棉花中来源于A 基因(亚)组的CRPK1 蛋白聚在一类,而来源于D 基因(亚) 组的CRPK1 蛋白聚在一类; 棉花中6 个CRPK1 蛋白与棉花近源种可可(Tc10v2_t005900) 及番茄 (Solyc09g007730)中CRPK1 蛋白聚在一类,而禾本科植物水稻、小麦和玉米中的CRPK1 分别聚在一起。 这些结果说明CRPK1 蛋白在进化上高度保守, 推测其在植物的生长发育中发挥着广泛而重要的作用。
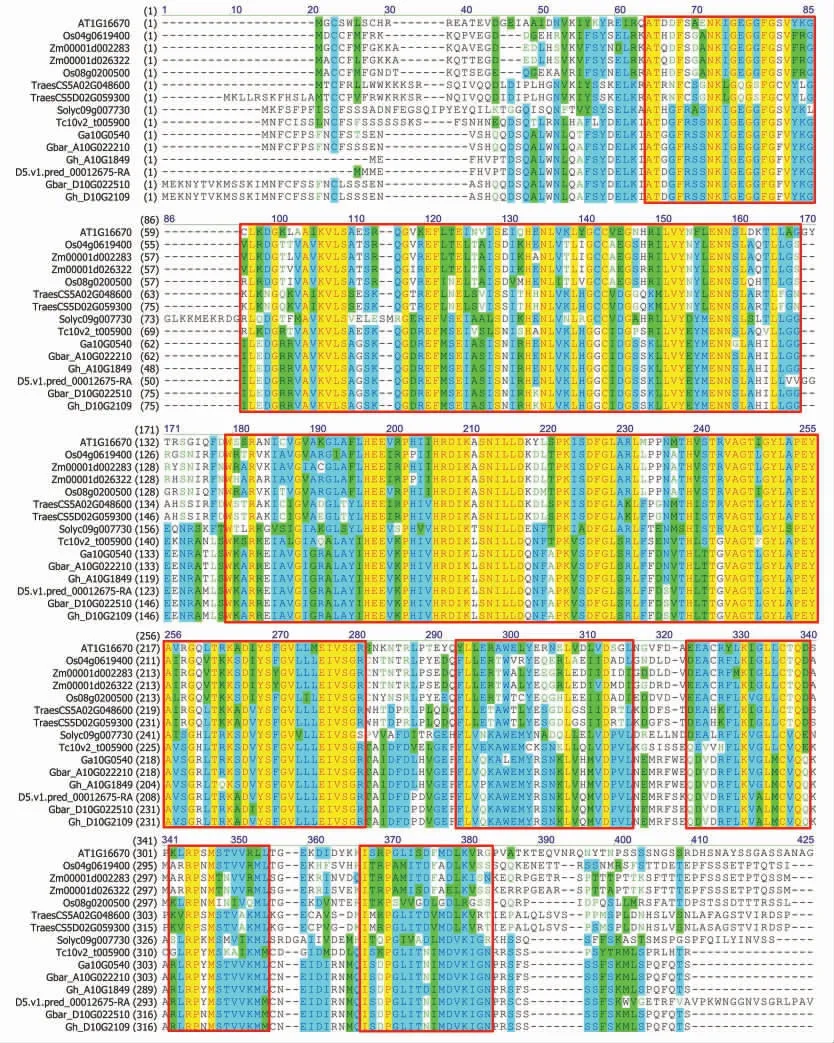
图2 多序列比对分析CRPK1 蛋白的保守结构域Fig. 2 Multiple sequence alignment of CRPK1 with its homologs and conserved domains analysis
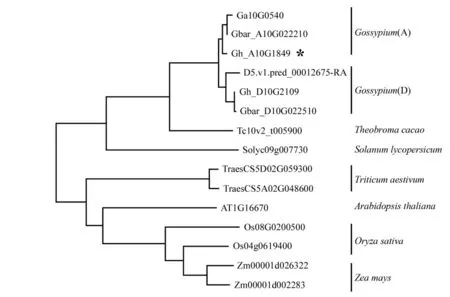
图3 GhCRPK1 蛋白与其同源蛋白的系统进化分析Fig. 3 Phylogenetic analysis of GhCRPK1 protein and its homologs from 9 species
2.3 GhCRPK1 蛋白的亚细胞定位分析
GhCRPK1 蛋白属于RLK 家族中的RLCK亚家族,一般定位在细胞膜。 为了确定GhCRPK1蛋白在细胞中的定位情况, 在烟草中观察融合蛋白的荧光分布, 对照组pCaMV35S∷GFP 载体的荧光信号分布在细胞膜和细胞核中,而融合蛋白载体pCaMV35S∷GhCRPK1-GFP 的荧光信号只分布在细胞膜上(图4),说明GhCRPK1 与传统的受体激酶类似,是一个膜定位蛋白。
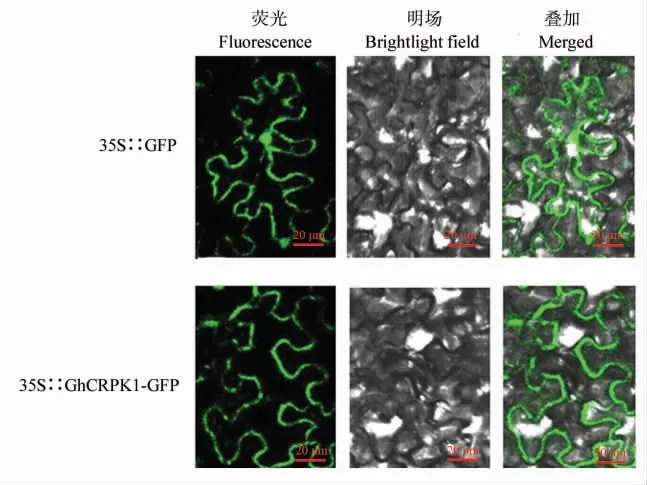
图4 GhCRPK1 蛋白的亚细胞亚定位Fig. 4 Subcellular location of GhCRPK1
2.4 GhCRPK1 基因的功能分析
为了进一步研究GhCRPK1基因的功能,构建pCaMV35S∷GhCRPK1-His 过表达载体,并转化到拟南芥中, 得到稳定表达GhCRPK1-His 的T3代转基因拟南芥植株。在体式显微镜下观察野生型(Wild type,WT)拟南芥、过表达GhCRPK1-His 拟南芥及拟南芥crpk1突变体莲座叶的表皮毛长度(图5)。通过观察和统计发现,野生型拟南芥的表皮毛平均长度为282.40 μm, 过表达GhCRPK1拟南芥的表皮毛的平均长度为340.60 μm, 而crpk1突变体的表皮毛的平均长度为263.60 μm(表1)。表明过表达GhCRPK1基因可促进拟南芥表皮毛的伸长; 拟南芥CRPK1基因突变会抑制拟南芥表皮毛的生长,导致表皮毛变短(图5)。 同时观察了3 个材料莲座叶表皮毛的密度,发现3 种材料表皮毛的密度没有显著差异(表1)。 这些结果说明,GhCRPK1正向调控拟南芥表皮毛的伸长,而不影响拟南芥表皮毛的密度。

表1 拟南芥莲座叶表皮毛长度和密度Table 1 The trichome length and density of rosette leaf in transgenic Arabidopsis

图5 野生型和转基因拟南芥表皮毛的表型Fig. 5 Phenotype of of trichomes in wild type and transgenic Arabidopsis
为了进一步分析GhCRPK1 蛋白发挥功能的分子机制,以GhCRPK1 为诱饵,在棉花纤维发育起始时期的蛋白表达文库中筛选到1 个与GhCRPK1 相互作用的蛋白,对应基因为Gh_D08G0057。功能注释发现,Gh_D08G0057基因属于HSP90 家族。 为了确定GhCRPK1 与Gh_D08G0057 的互作关系,我们在酵母中进行了点对点的互作验证,首先在酵母中对pGADT7-Gh_D08G0057 载体进行转录自激活检测, 发现Gh_D08G0057 蛋白在酵母中没有转录自激活活性;分别将pGADT7-Gh_D08G0057 和pGBKT7-GhCRPK1 共转化酵母后,发现只有pGBKT7-GhCRPK1 和pGADT7-Gh_D08G0057 质粒共转化的酵母能够在四缺培养基中长出菌斑(图6),说明GhCRPK1 与Gh_D08G0057 蛋白在酵母中能够发生相互作用。

图6 GhCRPK1 与Gh_D08G0057 蛋白在酵母中互作分析Fig. 6 Yeast two-hybrid interaction analysis of GhCRPK1 and Gh_D08G0057
3 讨论
CRPK1 是1 个冷响应蛋白激酶,属于类受体蛋白激酶。在拟南芥中,CRPK1 蛋白位于细胞膜,接收到外界环境传来的低温信号时,激酶活性被激活进而磷酸化14-3-3s 蛋白,促使14-3-3s 在细胞核中的积累, 进而调控低温相关转录因子CBF1 和CBF3 的蛋白活性水平, 激活下游冷响应相关基因的表达,调控植物对低温的响应[23-26]。但是拟南芥CRPK1 缺少典型的跨膜结构域,无法直接识别胞外信号, 因此CRPK1 识别胞外冷信号需要其它蛋白的协助。 拟南芥热激蛋白HSP90 也参与细胞冷胁迫反应介导的细胞死亡及防御反应[27]。热激蛋白在代谢较快、蛋白质大量合成的细胞中含量较多,具有稳定蛋白质或辅助蛋白质完成折叠的作用,在蛋白质复合物组装过程中保护蛋白质并防止蛋白质聚集和分解,同时对错误折叠的蛋白质进行降解[28]。 通过棉花全基因组关联分析数据, 在4 个环境中均能检测到GhCRPK1与纤维长度高度相关,并且GhCRPK1在开花当天、开花后1 d 和3 d 均高水平表达,与棉花纤维发育起始时期一致, 说明GhCRPK1可能参与了棉花纤维发育的起始。 将该基因导入拟南芥中过量表达后,能显著增加拟南芥表皮毛的长度,而拟南芥中的crpk1突变体的表皮毛变短,拟南芥的表皮毛和棉花纤维发育有着相似之处,表明GhCRPK1 在棉花纤维发育过程中发挥着重要的作用。 此外酵母双杂交试验表明GhCRPK1与热激蛋白Gh_D08G0057 相互作用, 可能参与表皮毛的调控。 陆地棉中的GhCRPK1 与拟南芥中的CRPK1 蛋白具有较高的同源性, 它们均没有明显的跨膜结构域,但均定位在细胞膜上发挥作用。 推测在棉花纤维发育过程中,由细胞表面的特定受体识别到环境中信号分子, 激活GhCRPK1 的激酶活性,进而诱发纤维发育或表皮毛发育相关基因的表达, 促进纤维或表皮毛的伸长。Gh_D08G0057 与GhCRPK1 相互作用,可能是辅助GhCRPK1 折叠形成正确的空间构象, 调节GhCRPK1 在细胞膜上的激酶活性以发挥功能。
4 结论
本研究克隆了陆地棉的受体激酶基因GhCRPK1,该基因广泛存在于高等植物中,其激酶结构域在不同的物种中同源性较高, 推测CRPK1 在不同的植物中具有相似的功能。 亚细胞定位发现GhCRPK1 定位在细胞膜上, 在拟南芥中稳定表达GhCRPK1基因能促进拟南芥表皮毛的伸长, 而拟南芥中的同源基因突变体crpk1表皮毛变短。 酵母双杂交试验表明GhCRPK1 与热激蛋白Gh_D08G0057 相互作用。 本文发现GhCRPK1 在调控拟南芥表皮毛发育过程中发挥着重要的作用,为探究其调控棉花纤维发育的功能提供了理论基础。
附表:
详见本刊网站(http://journal.cricaas.com.cn)本文网页版。
附表1 本研究中所用到的引物序列
Table S1 The sequence of primers used in this study
