三细胞紧密连接蛋白tricellulin表达与功能调控的研究进展*
2021-01-06毛祥娣杨泽希
毛祥娣,杨泽希,丛 馨△
(1北京大学基础医学院生理学与病理生理学系,2北京大学医学部临床医学院,北京 100191)
紧密连接(tight junction,TJ)是由多个蛋白构成的大分子复合物,广泛分布在所有上皮细胞和内皮细胞的顶侧膜区域。TJ通过调节水、离子和大、小分子物质经旁细胞途径的转运以及限制细胞膜脂质和蛋白的自由流动,发挥着重要的屏障和栅栏功能。狭义的TJ 复合物主要包括跨膜蛋白,如密封蛋白(claudin)家族、闭合蛋白(occludin)和连接黏附分子(junctional adhesion molecules,JAMs)等,以及胞浆蛋白,如闭锁小带蛋白(zonula occludens,ZO)和扣带素(cingulin)等[1];广义的TJ 复合物还包括与调控TJ相关的信号调控蛋白,如G 蛋白、非典型蛋白激酶C(atypical protein kinase C)和蛋白磷酸酶(protein phosphatase)等,以及转录因子,如激活蛋白1(acti⁃vator protein-1,AP-1)和ZO-1 相关核酸结合蛋白(ZO-1-associated nucleic acid-binding protein,ZONAB)等。
tricellulin 是第一个被发现的主要表达在三个相邻细胞接头处的TJ 跨膜蛋白。上述提到的TJ 跨膜蛋白主要分布在两个细胞间侧膜,被称为两细胞紧密连接(bicellular tight junction,bTJ),而tricellulin 与它们不同,因其分布特点而被称为三细胞紧密连接(tricellular tight junction,tTJ)。已有研究显示,tri⁃cellulin在封闭大分子物质经旁细胞途径的转运中发挥了决定性作用,并且对维持tTJ 和bTJ 结构的完整性至关重要。本文将从tricellulin 的结构、表达和分布、功能与疾病以及相关的调控机制几个方面进行综述。
1 tricellulin的结构
早在1969 年,Staehelin 等利用冰冻蚀刻技术观察到2 个相邻细胞间的TJ“拉链”在到达与第3 个细胞接触时变为垂直向基底方向延伸,因此在三细胞接头处并不汇聚,而是断开的。1973年,Staehelin在电镜下发现在三细胞接头处存在3 对与顶膜垂直的独立TJ 链,从而围成一个“中心管”(图1)。随后的30多年,研究者检测并深入研究了30多种bTJ分子,但受检测技术的限制一直未见tTJ 分子被报道。直到2005 年,Ikenouchi 等[2]首次发现并命名了tTJ 蛋白tricellulin。后来,研究者又证实,可调控tricellulin分布的angulin家族亦属于tTJ蛋白。
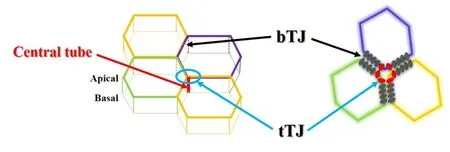
Figure 1.The structure diagram of tricellular and bicellular tight junctions.At tricellular contacts,there are 3 pairs of independent tight junction chains perpendicu⁃lar to the apical membrane,thus forming a "central tube".bTJ:bicellular tight junction;tTJ:tricellular tight junction.图1 三细胞紧密连接和两细胞紧密连接的结构示意图
tricellulin 是由位于染色体5q13.2 上的Marveld2基因编码,与常染色体隐性非综合性遗传性耳聋49位基因(autosomal recessive non-syndromic deafness gene 49,DFNB49)位点相连[3]。在蛋白结构上,人tricellulin 是一个由558 个氨基酸残基构成的四次跨膜蛋白,分子量约为64 kD,其N 末端和C 末端均位于胞浆内。由于共同具有一个保守的中央MARVEL域(MAL and related proteins for vesicle trafficking and membrane link),tricellullin(又称MARVELD2)、oc⁃cludin(又称MARVELD1)及MARVELD3 共同属于紧密连接相关的MARVEL 家族蛋白(tight junctionassociated MARVEL protein,TAMP)[4-5]。其中,tri⁃cellulin 和occludin的结构最为相似,均包含一个C末端的卷曲螺旋(coiled-coil,CC)结构域,该结构域与RNA 聚合酶II 转录因子ELL 家族的C 末端有很高的同源性[6]。已有研究表明,tricellulin 的C 末端在决定其在细胞膜上的定位中发挥重要作用,如其CC 结构域内氨基酸发生突变,可破坏它与ZO-1 的相互作用造成tricellulin 的定位异常,从而引起前庭和耳蜗旁细胞屏障功能障碍,最终导致听力损伤[7-8]。
2 tricellulin的表达和分布
正常的表达量和正确的分布是tricellulin 发挥屏障功能的前提。尽管tricellulin 是一个近年来新发现的tTJ 分子,但其在多种组织和器官中的表达和分布特点已相继被报道。
2.1 tricellulin 的表达 tricellulin 存在于哺乳动物多种上皮组织中,如小肠、胃、肾、肺、肝、睾丸、皮肤、前庭和耳蜗等,以及上皮细胞系,如肠上皮HT-29/B6、Caco-2 和T84 细胞、肾上皮MDCK I、MDCK II 和M-1 细胞以及乳腺上皮EpH4 细胞等[9-12]。Tricellulin还在脑和视网膜的微血管内皮以及脑微血管内皮细胞系(brain microvascular endothelial cells,BMECs)中存在表达[13]。据此推测,tricellulin 可广泛地表达于全身器官和组织的上皮及内皮细胞。
值得注意的是,在人单核细胞系THP-1 以及成熟的树突状细胞中也检测到tricellulin 的mRNA 表达,提示tricellulin 可能参与了树突状细胞与上皮细胞之间紧密连接样结构的形成,从而帮助维持上皮屏障的完整性[14]。此外,作为脑中常驻的重要免疫细胞的小神经胶质细胞中也发现tricellulin mRNA 和蛋白的表达[15],再次提示除了构成上皮和内皮屏障外,tricellulin 在免疫细胞中的表达及意义也值得深入研究。
2.2 tricellulin 的分布 一直以来,tricellulin 如何特异性地表达在三细胞接头处是很多学者关注的一个重要问题。目前研究认为,tricellulin 不能自行定位在三细胞间,需要通过与同为tTJ的angulin家族分子相互作用从而被招募到三细胞接头处[16-17]。angu⁃lin家族(angulin-1~3)是I型跨膜蛋白,具有位于细胞外的N 末端免疫球蛋白样结构域(immunoglobulinlike domain)和位于胞浆内的C末端。
angulin-1 又称脂解激活脂蛋白受体(lipolysis stimulated lipoprotein receptor,LSR)。在特异性敲低angulin-1的小鼠乳腺上皮EpH4细胞中,tricellulin不能很好地定位在tTJ 处,而是均匀分布在两细胞间,呈现与bTJs相似的分布模式,表明angulin-1对tricel⁃lulin 的分布具有重要的招募作用[18]。进一步研究证实,angulin-1蛋白易发生磷酸化修饰,在人类和小鼠中已检测到至少30 个磷酸化位点[19]。angulin-1 的C末端发生磷酸化后可与tricellulin 相互作用,从而招募tricellulin 到三细胞间[18-21]。angulin-2 又称含免疫球蛋白样结构域受体1(immunoglobulin-like domain containing receptor 1,ILDR1)。在Ildr1基因敲除小鼠的内耳上皮中,tricellulin 仍可定位在tTJ 处,且能较好地维持bTJ链与中心管的密封元件的连接,这可能与此时angulin-1 的补偿作用有关[22]。此外,angu⁃lin-2 的细胞外域可能在tTJ 处形成同源三聚体结构,然而其具体机制尚不清楚[23]。angulin-3 又称IL⁃DR2,与angulin-1 和-2 相比其招募tricellulin 的作用最弱[18],目前研究甚少。
除了angulins 分子的作用外,敲减occludin 也会导致tricellulin 重分布至两细胞间,部分补偿了此时occludin 的缺失[24]。在仅过表达tricellulin 的犬肾上皮MDCK II 细胞中,tricellulin 可同时分布在bTJ 和tTJ 处;而如果共同过表达tricellulin 和occludin,tri⁃cellulin 则仅分布在tTJ 处。这表明bTJs 分子的存在对tricellulin 在三细胞接头处的定位也是一个重要的影响因素。
3 tricellulin的功能及在疾病中的作用
3.1 tricellulin的功能 与bTJ所构成的两细胞侧膜间隙相比,tTJ 所组成的“中心管”具有更大的孔径(直径约10 nm),因此有学者提出tTJs 可能与一些大分子物质经旁细胞途径的转运有关,而bTJs 则与离子和小分子物质的转运有关。在MDCK 细胞中,过表达tricellulin 可降低大分子物质(分子量>4 kD)的通透性,但不影响离子转运[25],这与上述推测相符。但是,tricellulin 究竟是否影响离子转运仍存在争议。例如,在内耳中,tricellulin 主要表达在耳蜗和前庭上皮中,参与构成不同区室间的屏障;当tricellulin 基因发生突变时,屏障受到破坏,导致不同区室的淋巴液中离子的通透性增大,最终造成不同程度的听力损伤[26]。在EpH4 细胞中,敲低tricellulin 可同时影响tTJs和bTJs的形成,引起离子的通透性增加,提示tri⁃cellulin可通过影响bTJs结构的完整性进而间接调控离子转运[2]。此外,在MDCK 细胞中,tricellulin 还可作为氧化还原反应感受器在缺氧和还原剂的作用下发生由三细胞间向两细胞间的重分布,代偿了此时occludin的缺失,提示tricellulin在bTJs表达缺失时可发挥重要的屏障补偿作用[27]。这些研究表明,tricel⁃lulin 在调控旁细胞途径的通透性、尤其是在大分子物质的转运中发挥关键作用,tricellulin 的异常可参与一些上皮或内皮屏障功能障碍性疾病的发生发展;此外,tricellulin 对bTJ 结构和功能的影响及相互作用也值得关注。
3.2 tricellulin在疾病中的作用
3.2.1 非综合性遗传性耳聋 目前,Marveld2被认为是人类和小鼠常染色体隐性非综合性遗传性耳聋DFNB49 的致病基因[28]。tricellulin 基因敲除小鼠也表现出与耳蜗毛细胞退化有关的早发性渐进性听力损伤[29]。
tricellulin 的C 末端是受angulin 招募所必需的,一旦缺失或突变将导致tricellulin 无法定位在tTJ处,这被认为是非综合性遗传性耳聋的一个致病原因[18,30]。但除了耳聋这一症状外,患者没有出现其他明显的表型,这可能是因为内耳上皮缺失tricellu⁃lin 易产生较为严重的屏障功能障碍,而在其他器官中其他TJ 蛋白可能起到补偿作用有关。研究证实,tricellulin 第497 位精氨酸位点基因突变(R497*)能够过早终止蛋白质的形成,从而导致C 末端ELL 结构域的丧失和tricellulin 的错误定位,因此tricellulinR497*敲入小鼠呈现出与基因敲除小鼠相似的表型,由此证实C 末端位点突变所导致的tricellulin 错误定位可消除tricellulin 的屏障功能[30]。此外,occludin基因敲除小鼠也出现听力丧失,且在耳蜗毛细胞中发现tricellulin 的定位异常,这与前面提及的当occludin缺失时tricellulin 可发生从tTJ 向bTJ 处重分布的代偿作用有关,因而引起上皮屏障功能的丧失和毛细胞的凋亡,也再次表明tricellulin 在维持正常听力中起着重要的作用[18]。
3.2.2 炎症性肠病 在两种经典的炎症性肠病即溃疡性结肠炎和克罗恩病患者的肠上皮标本中检测发现,tricellulin 的蛋白表达水平仅在溃疡性结肠炎中显著下调,而在克罗恩病中不变[31]。在人结肠上皮HT/B6 细胞中,通过一种可视化实时观察方法显示只经旁细胞途径转运的大分子物质(分子量为10 kD 的右旋糖苷)主要是经由tricellulin 介导的tTJ 途径而通透的。动物模型进一步证明,在溃疡性结肠炎发生的早期,tricellulin 表达的下调可能通过破坏tTJ 的紧密性,从而允许抗原等大分子物质通过,最终启动局部的免疫炎症反应并加重疾病的进程。由此作者推测,在炎症性肠病的早期阶段,针对上皮屏障功能的缺失以及大分子药物在体内不受控制的吸收等改变,tricellulin 可能是一个潜在的有效治疗靶点。
3.2.3 细菌感染性疾病 研究证实,一些致病菌可以通过攻击tTJ 来有效感染上皮细胞。例如,引起细菌性痢疾的志贺痢疾杆菌可侵入结肠上皮并在结肠细胞中繁殖,而这一过程需要以tricellulin 依赖的方式在tTJ 处形成伪足而实现。另外,有学者发现A 组链球菌可通过与tricellulin 的相互作用攻击由tTJ 介导的大分子物质转运途径,进而破坏上皮屏障,这或许可以解释A 组链球菌能够更深入地侵入至黏膜下层组织的作用机制[32]。由此可见,tricellulin 可作为细菌入侵的一个重要攻击靶点,如能较好地维持tri⁃cellulin的表达、分布和屏障功能,将有望为尽早阻断细菌感染性疾病的发生发展提供新思路。
3.2.4 肿瘤 已有很多研究表明,TJ的表达异常可破坏屏障功能,使细胞极性丧失,造成细胞间黏附力的降低和细胞-细胞连接的松弛,破坏上皮细胞的接触抑制,进而促进细胞的异常增殖,最终导致多种肿瘤的发生发展。研究证明tricellulin 表达的减少与肿瘤的进展密切相关。例如,在扁桃体鳞状细胞癌患者受累组织中,tricellulin 和claudin-7 的蛋白表达减少[33]。与正常肝细胞相比,肝癌患者组织中tricellu⁃lin 的表达显著减少[34]。肝胆管癌的预后不良,与tri⁃cellulin 的表达减少相关[35]。在胰腺导管腺癌中,tri⁃cellulin 的表达水平与癌细胞的分化程度相关,分化好的腺癌中tricellulin 高表达,但在分化差的腺癌中表达较少[10]。此外,在分化不良的胰腺癌患者组织标本中,tricellulin 发生由细胞膜向细胞核的重分布,而核内表达的tricellulin 可进一步通过丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)和蛋白激酶C(protein kinase C,PKC)信号通路增强胰腺癌细胞的增殖和迁移能力,因此tricellulin 的核定位在一定程度上可反映胰腺癌的恶性程度[36]。进一步在人胰腺腺泡上皮癌HPAC 细胞中,特异性敲低tricellulin 可抑制细胞迁移并降低上皮屏障功能[37]。以上研究表明,tricellulin 的表达水平与多种上皮源性肿瘤的发生发展有关,tricellulin 的低表达可能是肿瘤进程及预后的一个潜在标志分子。
3.2.5 血脑屏障与血视网膜屏障 尽管内皮细胞tTJ 的结构和功能特征不如上皮细胞的明显,但最近的研究已开始关注到tricellulin 在血脑屏障(bloodbrain barrier,BBB)和血视网膜屏障(blood-retinal barrier,BRB)中的重要性。BBB 和BRB 的维持对神经元的稳态、局部营养物质的运输以及阻止炎性细胞和介质的侵入等生理功能有重要意义。研究显示,tricellulin 在脑和视网膜微血管内皮细胞的三细胞接头处存在表达[13]。angulin-1 基因敲除小鼠的脑微血管内皮细胞更容易渗透小分子物质(分子量<450 Da),而在多发性硬化和脑卒中等BBB 受损的动物模型中亦发现脑微血管内皮细胞中angulin-1 的表达显著下降,由于angulin-1 具有招募tricellulin 定位到tTJ处的作用,因此推测上述脑血管疾病时BBB 障碍的发生可能与tricellulin 有关,但具体机制尚在研究中[38]。
另外,有研究向小鼠尾静脉注射一种tTJ 的粘合剂——angubindin-1,发现它可破坏angulin-1 对tri⁃cellulin 的招募作用进而抑制tricellulin 的屏障功能,从而利于将反义寡核苷酸穿透BBB 转运到中枢神经系统使目标RNA 沉默,由此实现对中枢神经系统性疾病的靶向性治疗效果[39]。因此,寻找合适的能特异性调控tricellulin 开放和关闭的药物,将为临床上一些治疗性药物更好地穿透BBB和BRB等血管内皮屏障到达特定器官和组织提供新策略。
4 tricellulin相关的调控机制
尽管已有研究表明tricellulin 在多种器官和组织中广泛表达,且与多种疾病的发生发展也有密切关系,但目前关于tricellulin 受哪些信号分子及转录因子的调控及具体机制的研究仍十分有限。以下是近几年报道较多的可能调控tricellulin 的几个机制(图2)。
4.1 促炎因子白细胞介素13(interleukin-13,IL-13)作用通路 现已知多种促炎因子,包括肿瘤坏死因子α(tumor necrosis factor-α,TNF-α)、干扰素γ(interferon-γ,IFN-γ)和IL-13等,可通过改变TJ蛋白的表达、分布和功能进而影响上皮或内皮的屏障功能。
前面提及有学者在溃疡性结肠炎患者标本中证实tricellulin 的蛋白表达降低,经进一步检测显示这与在溃疡性结肠炎的发生发展中起关键始动作用的促炎因子IL-13 有关[31]。IL-13 可选择性激活IL-13受体α2(IL-13 receptor α2,IL-13Rα2),继而活化MAPK家族中的细胞外信号调节蛋白激酶1/2(extra⁃cellular signal-regulated kinase 1/2,ERK1/2)和c-Jun N 末端蛋白激酶(c-Jun N-terminal kinase,JNK),通过引起转录因子AP-1 入核、与tricellulin 启动子区结合下调tricellulin 的mRNA 和蛋白表达,从而增加大分子物质经tTJ 途径的通透性。这表明,IL-13 可通过IL-13Rα2/MAPK(ERK1/2 和JNK)/AP-1 信号通路下调tricellulin 的表达,从而造成大分子物质如抗原经三细胞间进入肠黏膜上皮产生局部免疫炎症反应。该作用机制的发现为早期治疗溃疡性结肠炎提供了潜在干预靶点,也丰富了我们对tricellulin 表达相关的信号调控机制的认识。
4.2 JNK/NF-κB信号通路 JNK是一种应激活化蛋白激酶,属于丝/苏氨酸蛋白激酶家族,其在细胞增殖、分化及凋亡等过程中发挥重要作用[40-41]。最近的研究发现,JNK 可参与调节不同上皮细胞中TJ 的完整性,包括对tricellulin 表达和功能的调控[42-43]。JNK的激活对人角质形成细胞和结肠上皮细胞的贴壁和TJ的开放至关重要[44-45]。在人胰腺HPAC 中,分别给予促炎因子IL-1β、TNF-α 和IL-1α 均可显著增加tri⁃cellulin 在tTJ 处的表达水平,增强上皮的屏障功能。进一步机制研究证实,JNK 是通过活化下游NF-κB信号通路来实现对tricellulin 在转录水平上表达的调控作用的[46]。以上结果提示,促炎因子可通过激活JNK/NF-κB 信号通路来上调tricellulin 的mRNA 和蛋白表达,而tricellulin 在tTJ处的表达增加以及相应的屏障功能增强可以阻断局部炎症反应,在炎症反应发生起始阶段发挥着一定的代偿性保护作用。
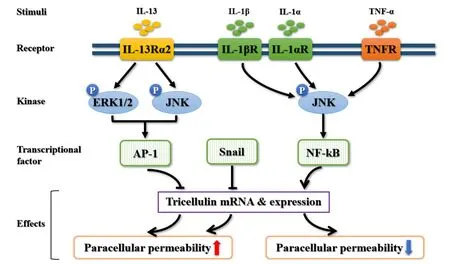
Figure 2.The related mechanism involved in the regulation of tricellulin.Interleukin-13(IL-13)activates extracellular signal-regu⁃lated kinase 1/2(ERK1/2)and c-Jun N-terminal kinase(JNK)through interleukin-13 receptor α2(IL-13Rα2),and then reduces the expression of tricellulin by activator protein 1(AP-1),eventually leading to increased permeability of macromo⁃lecular substances.Proinflammatory factors,such as interleukin-1α/β(IL-1α/β)and tumor necrosis factor-α(TNF-α),can up-regulate the mRNA and protein expression of tricellulin by activating the JNK/NF-κB signaling pathway.Snail has a negative regulation of tricellulin expression and induces redistribution of the protein to cytoplasm from tricellular tight junc⁃tion.TNFR:tumor necrosis factor receptor;IL-1α/βR:interleukin-1α/β receptor;NF-κB:nuclear factor-κB.图2 tricellulin相关的调控机制
4.3 Snail家族转录因子 上皮-间充质转化(epithe⁃lial-mesenchymal transition,EMT)的特征性表现是细胞间连接的减少和细胞极性的丧失,这同样也是TJ发生障碍的标志性改变。很多研究已证实,EMT 在上皮细胞来源的恶性肿瘤的转化及进程中发挥了重要的作用,而Snail 信号通路是诱导EMT 发生的经典信号途径之一。Snail 家族是一个转录因子家族,包括Snail 和Slug,通过引起EMT 和负性调节claudin 及occludin 等TJ 和黏附连接分子E-cadherin 的表达,在胚胎发育、组织重建和肿瘤进展中起关键作用[47-49]。值得注意的是,tricellulin 的发现正是通过在过表达Snail 的Eph4 细胞中采用高密度寡核苷酸芯片所筛选出来的,因而Snail 家族分子对tricellulin 的mRNA表达的调控作用是一个非常明确的机制[2]。此外,在胃癌患者标本中的研究发现,Snail诱导的EMT 对tricellulin 的表达有负调节作用[50]。在过表达Snail的人胃癌HSC-45细胞中,tricellulin的蛋白表达水平显著降低,且发生由tTJ 处向细胞浆的重分布[50]。因此,除上述提及的AP-1 和NF-κB 外,Snail 家族是另一类可参与调控tricellulin 表达的重要转录因子,其在多种上皮中对tricellulin 表达的负性调控作用不容忽视。
5 结语
综上所述,tricellulin 作为tTJ 的重要组成成分,是影响上皮和内皮TJ 屏障功能、特别是对大分子物质的通透性的关键蛋白之一。尽管目前对tricellulin的认识仍处于起步阶段,其与细胞内、外其他物质之间的相互作用关系、调节机制及定位表达等仍未完全明了,但从tricellulin 的角度研究疾病,尤其是在耳聋和炎症性疾病等方面的研究已成为热点。因此,深入研究tricellulin 的功能及调节机制,了解其与疾病之间的关系,将加深对TJ 相关疾病发生发展的理解并也可为治疗相应的疾病提供潜在干预靶点。
