文心兰响应软腐病菌胁迫的生理生化分析
2020-09-26吴秋桢林争春倪珊珊赖钟雄
吴秋桢 林争春 倪珊珊 赖钟雄


摘 要:通过细菌性软腐病菌液针刺接种文心兰‘小樱桃假鳞茎,分别在0、8、16、24、32 h时取假鳞茎上第一片叶,测定其叶绿素荧光特性[最大光合效率Fv/Fm、实际光合效率Y(Ⅱ)、调节性能量耗散的量子产额Y(NPQ)、非调节性能量耗散的量子产额Y(NO)],超氧化物歧化酶(SOD),过氧化物酶(POD),过氧化氢酶(CAT),多酚氧化酶(PPO)的酶活性,丙二醛(MDA),木質素及可溶性糖含量的变化。结果表明,随着侵染时间的延长,Fv/Fm、Y(Ⅱ)、Y(NPQ)逐渐下降,Y(NO)逐渐升高,光合系统受损;PPO活性及木质素含量逐渐下降;SOD、POD活性逐渐升高;CAT活性与可溶性糖含量先上升后下降;MDA含量在0~24 h逐渐下降,24 h后趋于平缓。探明文心兰感染软腐病后其抗性机制,建立抗性鉴定的生理方法。
关键词:文心兰;软腐病;生理生化指标;叶绿素荧光特性;木质素
中图分类号:S682.31 文献标识码:A
Abstract: The pseudobulb of Sherry Baby, Oncidium was infected by bacterial soft rot bacteria infection. The first leaf on the pseudobulb was sampled 0, 8, 16, 24, 32 h, after infection to determine the chlorophyll fluorescence properties [Fv/Fm, Y(Ⅱ), Y(NPQ), Y(NO)], superoxide dismutase(SOD), peroxidase(POD), catalase(CAT), polyphenol oxidase(PPO) activity, malondialdehyde (MDA), lignin, and the change of soluble sugar content. The results showed that with the extension of infection time, Fv/Fm, Y(Ⅱ), Y(NPQ) gradually decline, Y(NO) increased, and the photosynthetic system was damaged. PPO activity and lignin content decreased gradually; SOD and POD activity increased gradually; CAT activity and soluble sugar content first increased and then decreased; MDA content decreased gradually from 0 to 24 h and then leveled off after 24 h. The study would heip to explore the resistance mechanism and establish the physiological method of resistance identification.
Keywords: Oncidium spp.; soft rot; physiological and biochemical indexes; chlorophyll fluorescence properties; lignin
DOI: 10.3969/j.issn.1000-2561.2020.08.015
文心兰(Oncidium spp.)又名舞女兰。其花枝长,花小而繁多,花姿优美,花色艳丽,是一种观赏价值较高的洋兰,被广泛用作切花材料和盆栽观赏。我国主要在台湾、福建、广东、云南、海南等地种植,研究表明其在15~25 ℃环境生长最适宜[1],而福建夏季高温多雨,正值文心兰切花产期,易受病原菌浸染,由欧文氏菌(Erwinia spp.)侵染而引起的软腐病(soft rot)是文心兰栽培生产最致命的病害之一,感病后植株迅速水渍化,后期常引起整株植株腐烂致死[2],危害严重且暂无有效的防治措施,越来越引起人们的关注。
植物感染软腐病后,会产生系列生理生化变化。白菜[3]、梨[4]等植物在感病后均呈现不同的生理状态,目前,学者对大花惠兰[5]、蝴蝶兰[6]等兰科植物响应软腐病胁迫进行相关研究,但未见有关文心兰感染软腐病生理变化的报道,因此,本实验以‘小樱桃文心兰(Oncidium cv. Sherry Baby)接种软腐病菌液,检测其叶绿素荧光特性的变化,SOD、POD、CAT、PPO活性,MDA、木质素和可溶性糖含量等,探明文心兰感染软腐病后其生理生化指标的变化规律,揭示侵染机制,为筛选文心兰抗软腐病植株及防治软腐病提供参考依据。
1 材料与方法
1.1 材料
材料由福建农林大学园艺植物生物工程研究所提供。选择1年生生长良好,且长势一致的‘小樱桃文心兰组培移栽苗。注射10 L的软腐病菌液(OD值为0.4)于文心兰假鳞茎,将植株套袋并放置在气候箱中,设置温度为36 ℃,湿度80%,光照强度为2000 lx和16 h光照、8 h黑暗循环处理。每组设3个对照,依次在0、8、16、24、32 h取样。处理结果如图1,取样后用液氮预冷处理并保存于80 ℃冰箱备用。
1.2 方法
1.2.1 叶绿素荧光参数测定 采用叶绿素荧光成像仪IMAGING-PAM(Germany, Walz),测定感染软腐病后最上部第一片叶片的叶绿素荧光参数,包括PSII最大光合效率(Fv/Fm)、实际光合效率[Y(Ⅱ)]、PSII调节性和非调节性能量耗散的量子产额[Y(NPQ)与Y(NO)]等指标。测定前对各处理的叶片暗适应遮光30 min,以确保PSII反应中心完全开放,各组处理设置2次重复,每样品测定5个区域。根据仪器说明书设定工作参数进行操作,系统运行结束后自动给出参数值,PSII最大光合效率通过公式Fv/Fm=(FmFo)/Fm计算获得。
1.2.2 相关生理指标测定 取文心兰叶片测定SOD、CAT、POD、PPO、MDA、木质素和可溶性糖等相关生理生化指标,3次重复。测定均采用苏州科铭生物技术有限公司的试剂盒,使用紫外可见分光光度计(上海元析仪器有限公司)测定相关波长下的吸光值,按照相关说明书进行操作和计算。
1.3 數据处理
试验数据分别采用Microsoft Excel、SPSS 17.0和GraphPad Prism 6.0软件进行统计与分析。
2 结果与分析
2.1 软腐病对文心兰叶绿素荧光参数的影响
2.1.1 文心兰感染软腐病前后Fv/Fm和Y(Ⅱ)的变化 从图2可知,侵染软腐病后,随处理时间的增加,文心兰叶片PSII的Fv/Fm和Y(Ⅱ)变化趋势基本一致,均呈下降趋势,0~8 h和16~24 h时下降幅度较小,分别比对照下降1.93%和12.75%,8~16 h时下降幅度最大,为对照的11.40%,24~32 h时Fv/Fm和Y(Ⅱ)下降幅度差别较大,Fv/Fm比对照下降17.79%,Y(Ⅱ)比对照降低21.91%。
2.1.2 文心兰感染软腐病前后Y(NPQ)与Y(NO)的变化 从图3可知,侵染软腐病后,随处理时间的增加,Y(NPQ)与Y(NO)的变化呈相反趋势。0~8 h时Y(NPQ)与Y(NO)变化缓慢,分别比对照降低和提高1.85%和7.40%;8~24 h时变化较大,Y(NPQ)降低11.55%,Y(NO)提高47.10%;24~36 h时Y(NPQ)与Y(NO)急剧下降和上升,Y(NPQ)下降至对照的22.78%,Y(NO)比对照提高了91.10%。
2.2 文心兰感染软腐病前后植物防御酶活性的变化
2.2.1 软腐病侵染对文心兰SOD活性的影响 从图4可知,侵染软腐病后,随处理时间的增加,文心兰叶片中SOD活性呈不断上升趋势,但在不同时期活性增幅有显著差异,处理0~8 h时SOD活性上升较缓,16~24 h时SOD活性上升同样较缓,但比0~8 h上升快,8~16 h和24~32 h时SOD活性显著提高。32 h后SOD活性为143.0783 U/g,比对照组20.1424 U/g提高7.1倍。
2.2.2 软腐病侵染对文心兰CAT活性的影响 从图5可知,侵染软腐病后,文心兰叶片0~8 h时CAT活性与对照组没有明显变化,8~16 h时CAT活性持续上升,且在16 h达到最大值,为481.380 nmol/(min·g),比对照提高26.58%,16~24 h时CAT活性显著下降,在24 h时活性与对照组相近,24~32 h时缓慢下降,此时CAT活性比对照组低10.92%。
2.2.3 软腐病侵染对文心兰POD活性的影响 从图6可知,侵染软腐病后,随着处理时间的增加,文心兰叶片POD活性呈不断显著上升的变化趋势,且增幅较稳定,8、16、24、32 h时POD活性分别比对照组增长21.43%、50.71%、85.71、114.29%。
2.2.4 软腐病侵染对文心兰PPO活性的影响 从图7可知,侵染软腐病后文心兰叶片中PPO活性随着处理时间的延长,总体呈下降趋势。在不同时期,下降幅度有所差异,0~8 h下降明显,比对照组降低18.25%,8~16 h呈平缓降低趋势,降幅为22.50%,16~24 h和24~32 h下降幅度相同,至32 h时比对照组降低53.33%。
2.3 软腐病侵染对文心兰MDA含量的影响
从图8可知,侵染软腐病后,0~16 h时文心兰叶片MDA含量缓慢下降,但MDA含量0~8 h时比8~16 h降幅略大,16~24 h时继续下降,且下降幅度显著,至24 h时MDA含量降至最低,24~32 h时无明显变化。
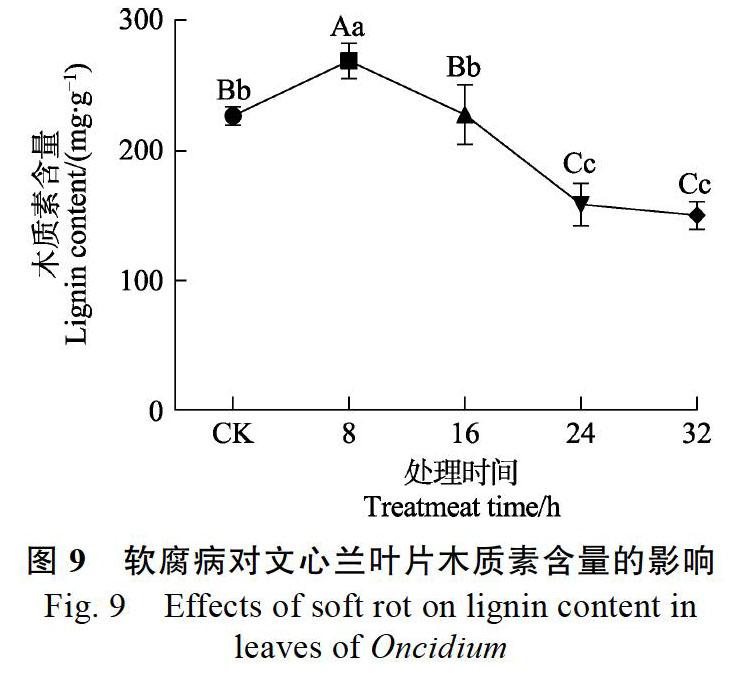
2.4 软腐病侵染对文心兰木质素含量的影响
从图9可知,侵染软腐病后,随着处理时间的延长,文心兰叶片木质素含量呈先增加后降低的趋势。0~8 h木质素含量上升,比对照组高18.73%,8 h后开始下降,至16 h时与对照含量相近,16~ 24 h下降显著,相比对照组低30.06%,24 h后变化不明显。
2.5 软腐病侵染对文心兰可溶性糖含量的影响
从图10可知,侵染软腐病后,文心兰叶片中可溶性糖含量呈先增加后下降的趋势,且上升与下降幅度明显。0~16 h显著增加,在16 h时达到最高水平,其含量为7.22 mg/g,16 h后开始下降,至32 h时比对照组低4.18%。
3 讨论
3.1 软腐病侵染后Fv/Fm和Y(Ⅱ)下降,叶绿体结构可能受到破坏
叶绿素对光能的吸收、传递以及耗散等过程都可以通过叶绿素荧光参数的变化反映,其是描述植物光合作用机理和光合生理状况的重要指标[7],在文心兰浅绿条纹突变体中最大光合效率(Fv/Fm)和实际光合效率[Y(Ⅱ)]均比正常植株下降,是由于叶绿体结构受到破坏[8]。而本研究中,文心兰叶片的最大光合效率(Fv/Fm)、实际光合效率[Y(Ⅱ)]和调节性能量耗散的量子产额[Y(NPQ)]在感染软腐病后均不断下降,而非调节性能量耗散的量子产额[Y(NO)]呈上升趋势,这与任少华等[9]、李丹等[10]的研究结果一致,这表明文心兰在感染软腐病后,PSII反应中心可能受到破坏,光合链上的电子传递轨道受阻,受光抑制作用。
3.2 软腐病侵染后期过氧化氢代谢失衡,自由基减少,防御酶系统受损
SOD、POD、CAT和PPO等作为防御酶系统,具有保护细胞免受损伤,对机体的氧化与抗氧化平衡起着至关重要的作用[7, 11]。SOD通过歧化反应清除自由基,生成过氧化氢,POD和CAT使其含量上升[12]。本研究中, SOD与POD随着感病时间的延长,活性不断提高,这与病毒感染百合[12]、枯萎病胁迫西瓜[13-14]、病菌胁迫香蕉[15]结果一致。CAT活性呈先上升,随后不断降低趋势,至低于对照组,表明感病后最初文心兰体内CAT活性增强,但这种变化可能无法满足清除持续产生的过氧化氢,导致过氧化氢累积,引起植株体内的过氧化氢代谢失衡,CAT活性不断下降[16]。文心兰感染软腐病后,自由基含量达到相对稳定,SOD清除自由基,生成过量的过氧化氢,POD与CAT又不断地清除过量的过氧化氢,以减轻对植物体的伤害[17]。PPO与植物的抗病性有关,通过与底物结合,合成醌类物质,构建防御系统[18]。本研究中,随着处理的延长,PPO活性不断下降,这与张晓南等[19]的研究结果不同,可能是由于病原菌浸染,文心兰腐烂,使其生理功能丧失所致。
3.3 软腐病侵染后期木质素合成受阻,细胞壁强度下降,导致软腐
MDA是膜脂过氧化的最终产物,其含量可作为逆境胁迫的重要指标[20-21]。本研究中,文心兰感染软腐病后,随着时间的延长,MDA含量不断降低,这可能是软腐病菌浸染细胞后,细胞膜体系遭到破坏,膜脂发生降解紊乱了植物细胞的生化平衡[22-23],这在真菌对百合耐热性[22]、木霉菌对黄瓜幼苗生长[24]的研究中有类似情况。木质素的合成与POD、PPO、PAL等多种酶有关,植物通过改变木质素的含量来调节细胞壁的强度,从而抵制病原菌入侵[20],本实验文心兰叶片中木质素在8 h时达到最大,随后开始下降,这与迟建国[25]和王乾等[26]的研究结果一致,病原菌浸染初期,木质素含量增加,从而提高细胞壁强度,抑制病原菌入侵。而一段处理时间后,相关合成酶降低,木质素合成受损,组织软腐。可溶性糖是渗透调节物质,帮助植物抵抗逆境,在浸染病原菌后,可溶性糖先增加后降低,表明在前期文心兰通过提高可溶性糖含量来抵御病原菌的入侵,随着处理时间延长,后期细胞被破坏,调节系统紊乱,可溶性糖显著下降。这与富贵竹感染茎腐病、大豆感染根腐病[27-28]等有类似情况。
综上所述,文心兰‘小樱桃品种在感染软腐病后,叶绿体结构受到破坏,光合作用受阻,防御酶系统遭到破坏,积累过量过氧化氢,自由基代谢失衡。同时,在感病后期,木質素合成受阻,细胞壁强度下降,进一步导致软腐。通过对叶绿素荧光参数及SOD、POD、CAT、PPO活性、丙二醛、木质素、可溶性糖含量的规律性变化,可作为文心兰抗病鉴定的关键指标,为培育抗病品种提供参考。
参考文献
钟士传. 蝴蝶兰软腐病防治试验[J]. 特产研究, 2005(3): 13-14.
辛海云. 文心兰栽培管理技术[J]. 中国园艺文摘, 2010, 26(10): 130-131.
张耀伟. 大白菜抗软腐病生理生化指标研究[D]. 哈尔滨: 东北农业大学, 2002.
王 倩. H2S延长梨果实及甘薯块根采后贮藏期的抗氧化机制研究[D]. 合肥: 合肥工业大学, 2012.
秦建彬, 陈俊杰, 魏翠华,等. 大花蕙兰软腐病防治药剂及防治适期的研究[J]. 农学学报, 2013, 3(3): 10-12, 38.
沈会芳, 蒲小明, 张景欣, 等. 蝴蝶兰细菌性软腐病绿色防治药剂筛选试验[J]. 广东农业科学, 2017, 44(6): 104-108.
王学亏, 黄见良. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2002: 134-137.
田韦韦, 王彩霞, 田 敏, 等. 文心兰浅绿条纹突变体的生理生化及叶绿素荧光特性研究[J]. 西北植物学报, 2015, 35(10): 2012-2017.
任少华, 李媛媛, 邹 军, 等. 高温胁迫对四种兰科植物叶绿素荧光参数的影响[J]. 北方园艺, 2017(12): 70-74.
李 丹, 黄玉吉, 朱 宁, 等. 三明野生蕉抗寒特性的形态学与生理生化研究[J]. 果树学报, 2019, 36(1): 76-85.
Zhu S, Ma B. Benzothiadiazole- or methyl jasmonate-induced resistance to Colletotrichum musae in harvested banana fruit is related to elevated defense enzyme activities[J]. Journal of Horticultural Science & Biotechnology, 2007, 82(4): 500-506.
李焕改. 百合主要病毒基因组的克隆及感病生理研究[D]. 大连: 大连理工大学, 2010.
陈 亮, 陈年来. 西瓜叶片防御酶活性与枯萎病抗性的关系[J]. 河南农业科学, 2019, 48(1): 77-83, 114.
孙正祥, 王 丰, 周 燚. 内生菌XG-1对西瓜枯萎病诱导抗性的研究[J]. 河南农业科学, 2013, 42(3): 71-75.
覃柳燕, 郭成林, 黄素梅. 棘孢木霉菌株PZ6对香蕉促生效应及枯萎病室内防效的影响[J]. 南方农业学报, 2017, 48(2): 277-283.
刘学庆, 王秀峰, 朴永吉. 蝴蝶兰不同品种耐冷特性的研究[J].园艺学报, 2007, 34(2): 425-430.
逯明辉, 宋 慧, 李晓明, 等. 冷害过程中黄瓜叶片SOD、CAT和POD活性的变化[J]. 西北植物学报, 2005(8): 1570-1573.
贺立红, 宾金华. 高等植物中的多酚氧化酶[J]. 植物生理学通讯, 2001(4): 340-345.
张晓南, 孙佩佩, 茅云翔, 等. 紫菜腐霉侵染对条斑紫菜叶状体防御性物质含量及保护性酶类活性的影响[J]. 中国海洋大学学报(自然科学版), 2015, 45(12): 57-64.
丁玉梅, 马龙海, 周晓罡等. 干旱胁迫下马铃薯叶片脯氨酸、丙二醛含量变化及与耐旱性的相关性分析[J]. 西南農业学报, 2013, 26(1): 106-110.
Shishatskaya E, Menzianova N, Zhila N, et al. Toxic effects of the fungicide tebuconazole on the root system of fusarium-infected wheat plants[J]. Plant Physiology and Biochemistry, 2018(132): 400-407.
邢红爽, 张 瑞, 郭绍霞. 高温胁迫下丛枝菌根真菌对百合耐热性的影响[J]. 青岛农业大学学报(自然科学版), 2018, 35(4): 258-264.
徐叶挺, 李 疆, 罗淑萍, 等. 低温胁迫下野生巴旦杏抗寒生理指标的测定[J]. 新疆农业大学学报, 2008, 31(4): 1-4.
高长敏, 马光恕, 廉 华, 等. 木霉菌对黄瓜幼苗生长和膜脂过氧化指标的影响及对枯萎病的防治效果[J]. 中国生物防治学报, 2018, 34(5): 762-770.
迟建国. 白腐菌对烟叶木质素含量的影响[J]. 贵州农业科学, 2013, 41(7): 138-139.
王 乾, 杨 亮, 胡艳敏, 等. 5个构树地方品种耐寒性状研究[J]. 北京农学院学报, 2019, 34(2): 94-96.
李 赤, 于 莉, 刘付东标, 等. 富贵竹中可溶性糖、蛋白质含量与细菌性茎腐病的关系[J]. 吉林农业大学学报, 2007(6): 620-622.
魏 崃, 王伟威, 李馨园, 等. 大豆抗腐霉根腐病的生理差异研究[J]. 大豆科学, 2017, 36(3): 425-429.
