波罗蜜叶绿素缺失突变体嫩茎转录组分析
2020-09-26郑李婷于旭东蔡泽坪罗佳佳吴繁花董俊娜曹佩娜
郑李婷 于旭东 蔡泽坪 罗佳佳 吴繁花 董俊娜 曹佩娜
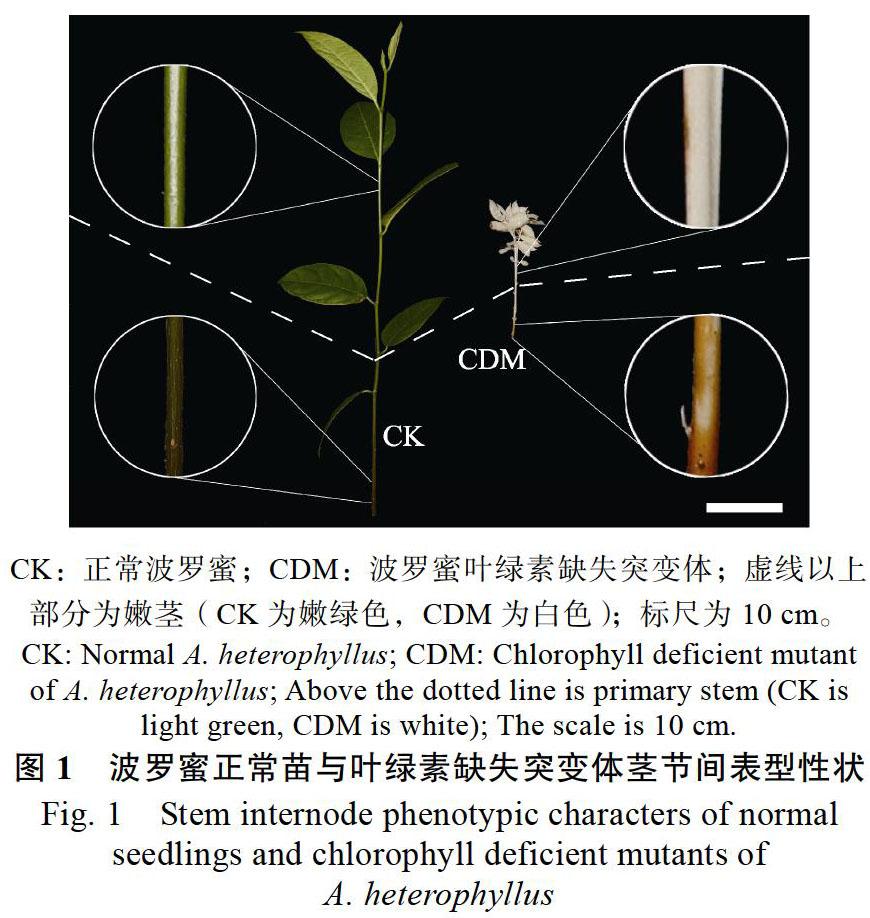
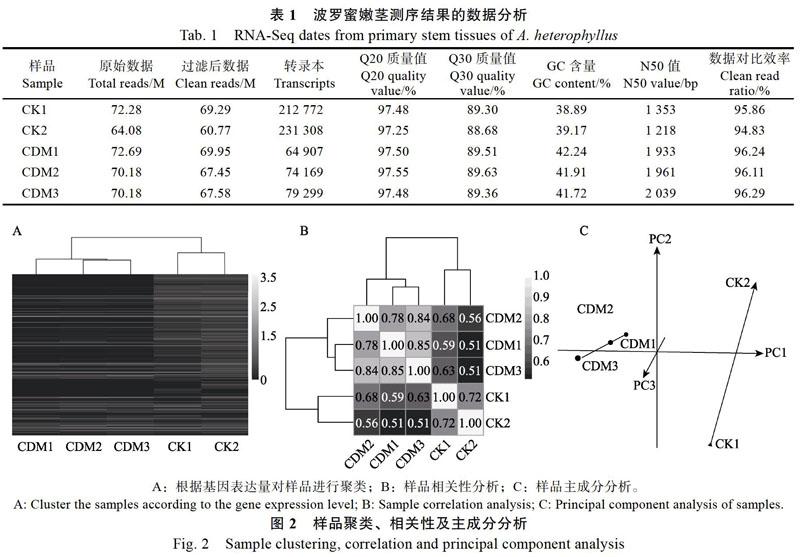
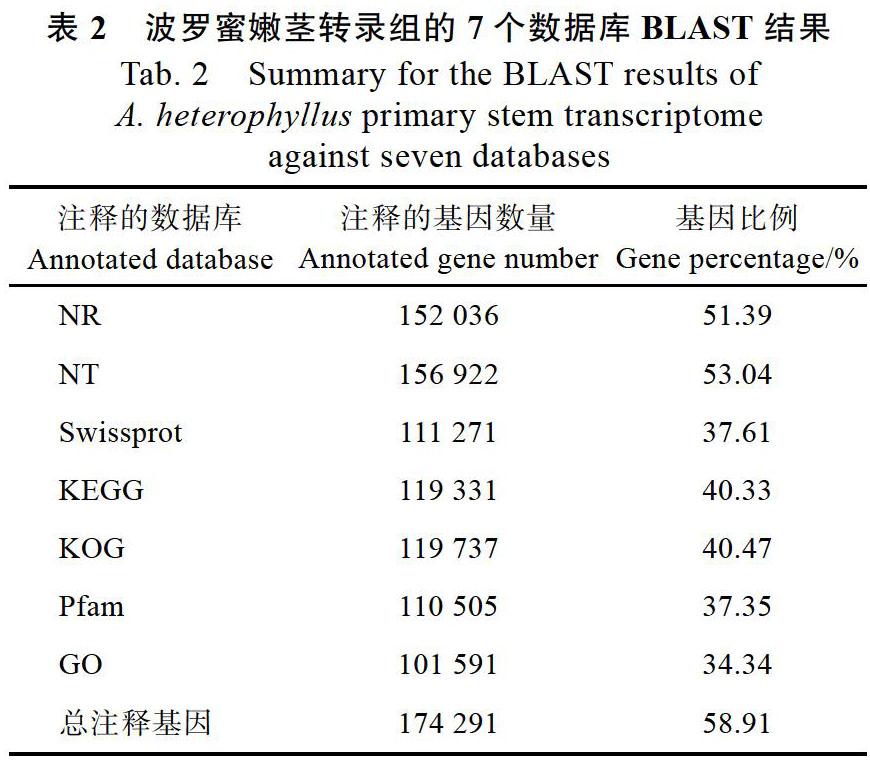
摘 要:为探究叶绿素合成缺失对波罗蜜嫩茎发育的影响,以波罗蜜叶绿素缺失突变体(chlorophyll deficient mutant, CDM)嫩茎为材料进行转录组分析;经de novo组装后获得295 869个Unigenes,使用NR、NT、Swissprot、KEGG、KOG、Pfam和GO数据库进行序列比对,共注释了174 291个Unigenes。过滤低丰度基因后,筛选出22 988个差异基因。与对照(CK)相比,CDM中上调基因有379个,下调基因有22 609个。GO分类结果表明,共有3712个基因获得注释,并将其划分为分子功能(molecular function)、细胞组分(cellular component)和生物学过程(biological process)三大类,共48个功能组;此外,有2080个Unigenes参与到19条KEGG通路上,其中在黄酮类生物合成途径中有30个Unigenes。该研究通过转录组测序,分析叶绿素合成缺失对波罗蜜嫩茎发育的影响,为木本植物茎发育的研究提供数据基础。
关键词:波罗蜜;叶绿素缺失突变体;嫩茎;转录组分析
中图分类号:S718.43 文献标识码:A
Abstract: To explore the influence of chlorophyll synthesis deficiency on the primary stem development of Artocarpus heterophyllus, the transcriptome of the primary stem of a chlorophyll deficient mutant (CDM) of A. heterophyllus was analyzed. After assembly by de novo, 295 869 Unigenes were obtained. NR, NT, Swissprot, KEGG, KOG, Pfam and GO databases were used for sequence alignment. A total of 174 291 Unigenes were annotated. After filtering the low abundance genes, 22 988 differentially expressed genes were selected. Compared with the control check (CK), there were 379 genes up-regulated in CDM and 22 609 genes down-regulated. GO classification results showed that 3712 genes were annotated and classified into three categories: molecular function, cellular component and biological process, with a total of 48 functional groups. In addition, 2080 Unigenes were involved in 19 KEGG pathways, 30 of which were involved in flavonoids biosynthesis pathways. Through transcriptome sequencing, this paper analyzed the effect of chlorophyll synthesis deficiency on the primary stem development of A. heterophyllus, which would provide data foundation for the study on the stem development of woody plants.
Keywords: Artocarpus heterophyllus; chlorophyll deficient mutant; primary stem; transcriptome analysis
DOI: 10.3969/j.issn.1000-2561.2020.08.002
茎上承枝叶下接根部,在植物生长过程中主要起运输和支撑的作用。茎的生长包括初生生长和次生生长。初生生长是顶端分生组织,经细胞分裂分化形成表皮、皮层和维管柱的过程[1]。国外学者通过对毛白杨(Populus tomentosa)各节间进行转录组测序,发现其茎主枝伸长区含特殊的碳水化合物活性酶(carbohydrate active enzymes, CAZymes)及扩展蛋白(expansins)家族成员基因。其中CAZymes中的糖基水解酶(glycoside hydrolases, GHs)小家族基因在茎伸长区显著性表达,部分成员可能参与初级细胞壁代谢(primary cell wall metabolish)、细胞壁疏松(cell wall loosening)和细胞伸长(cell elongation)[2]。Expansins是植物细胞扩张和疏松的关键调控因子[3-4]。而在毛白杨茎发现数个expansins基因在茎伸长区表达,可能参与初级细胞壁的合成和修饰[2]。三羧酸循環在毛白杨茎发育早期呈下降趋势,可能与丙酮酸脱氢酶(pyruvate dehydrogenase, PHD)、异柠檬酸脱氢酶(isocitrate dehydrogenase, ICDH)和琥珀酸脱氢酶(succinate dehydrogenase, SDH)蛋白丰度的降低有关[5]。
2.2 差异基因的筛选与分析
在CK和CDM中,平均表达量分别≤0.5的基因有70 218和94 226个,0.5~1的有59 403和23 361个,1~2的有77 073和20 733个,>2的有76 495和41 680个(图3A)。过滤平均表达量≤0.5的基因后,筛选出22 988个差异基因(图3B)。将其分为Ⅰ(CK中特有基因)、Ⅱ(CK与CDM共有基因在CDM中下调)、Ⅲ(CK与CDM共有基因在CDM中上调)和Ⅳ(CDM中特有基因)(图3C)4部分,每个部分转录因子(Transcription factors,TFs)各有41、163、7和3个,抗性基因(plant resistance genes,PRGs)各有87、193、20和3个(图3D)。通过WGCNA分析CK与CDM间差异基因的关联模式,结果显示所有差异基因属于一个共表达模块(图3E),且同组样品间特征向量基因表达趋于一致(图3F)。
2.3 差异基因GO、KEGG功能注释
2.3.1 GO功能注释 将22 988个差异基因进行GO功能注释和分析,可确定其重要的生物学功能。结果表明,共有3712个基因获得注释,并划分为48个功能组(图4)。因功能组的不同又可进一步分为三大主类:分子功能(molecular function)、细胞组分(cellular component)和生物学过程(biological process)。三类注释分别占79.85%、56.17%和46.82%,表示大部分差异基因与分子功能相关。分子功能注释的基因大部分集中在催化活性(catalytic activity),且与CK相比,CDM中120个基因上调,1787個基因下调,其中CDM特有的有16个。在生物学过程中,以细胞过程(cellular process)的基因数最多,且与CK相比,CDM中大部分差异基因下调,少数上调,其中CDM特有的有8个;在细胞组分中,膜(membrane)功能组拥有的基因数最多,且CDM相对于CK发生大量下调,少数上调,其中CDM特有的有6个。
2.3.2 KEGG功能注释 对差异基因进行KEGG途径分析,根据对结果中的代谢(metabolism)、遗传信息处理(genetic information process)、环境信息处理(environmental information processing)、细胞过程(cellular processes)以及生物系统(organismal systems)5组类目进行归类,有助于深入了解CK与CDM间差异基因所在的通路情况(图5)。结果表明,2 080个Unigenes被注释到19个条目,其中10个在Ⅰ、Ⅱ、Ⅲ和Ⅳ 4部分中均有出现。代谢类目注释的Unigenes最多,有1152个(55.38%),且以糖代谢(carbohydrate metabolism)注释的Unigenes最多,其中CK和CDM特有的分别有109和2个。而在遗传信息处理中,有698个(33.56%)Unigenes得到注释,翻译(translation)所含的Unigenes最多,且CDM相比于CK发生大量下调,少量上调,CDM特有的有2个。环境信息处理、细胞过程及组织系统所注释的Pathway较其他分类少,分别为2、1、1条。
2.4 GO富集分析
为进一步研究波罗蜜CDM嫩茎发育过程中基因的生物学功能,将差异基因进行GO富集分析(图6)。与CK相比,CDM有379个基因上调,22 609个基因下调。通过Q-value值分别筛选出20条最显著的条目。结果表明,在上调基因中,20个GO条目富集显著程度相同,而翻译(translation)和信号转导(signal transduction)是富集基因数较多的GO条目。在下调基因中,单加氧酶活性(monooxygenase activity)、多细胞生物发育(multicellular organism development)和作用于成对供体上的氧化还原酶活性,并结合或还原氧分子(oxidoreductase activity, acting on paired donors, with incorporation or reduction of molecular oxygen)3个GO条目最显著富集,而膜的组成成分(interal component of membrane)是富集基因最多的GO条目。
2.5 KEGG富集分析
通过KEGG Pathway富集分析,确定差异基因参与的最主要途径,在差异基因中找出显著富集的pathway(图7)。KEGG富集分析结果显示,脂肪酸降解(fatty acid degradation)、光合作用(photosynthesis)、苯丙氨酸代谢(phenylalanine metabolism)和色氨酸代谢(tryptophan metabolism)是上调基因和下调基因共有的富集代谢通路。在上调基因中,获得了5个氧化磷酸化(oxidative phosphorylation)途径的Unigenes,是富集基因最多的代谢通路。而在下调基因中,油菜素甾醇生物合成(brassinosteroid biosynthesis)和苯丙烷生物合成(phenylpropanoid biosynthesis)是最为显著富集的代谢通路。其次,二萜类生物合成(diterpenoid biosynthesis)、角质,琥珀和蜡的生物合成(cutin, suberine and wax biosynthesis)、黄酮类生物合成(flavonoid biosynthesis)和玉米素生物合成(zeatin biosynthesis)等是较为显著富集的通路,苯丙烷生物合成(phenylpropanoid biosynthesis)是富集基因最多的代谢通路。
3 讨论
近年来,随着转录组测序技术的发展,为波罗蜜分子生物学方面研究提供极大便利。学者通过对波罗蜜花被进行RNA-seq分析,探究其糖代谢相关基因差异表达的特点[24]。本研究通过转录组测序,分析了叶绿素合成缺失对波罗蜜嫩茎发育的影响。
波罗蜜是一种具有较高药用价值的热带水果,一直深受人们喜爱,也获得了众多学者青睐[25]。张福平等[26]研究了波罗蜜种子中黄酮类化合物的抗氧化性;国外学者测定了新鲜波罗蜜果实球茎中总黄酮类化合物及自由基清除活性在气调贮藏过程中的变化[27]。黄酮类化合物能清除大量自由基,此生理生化屏障在植物防御中发挥着重要作用[28]。本研究通过KEGG富集分析,获得了30个黄酮类生物合成途径的Unigenes,且均下调表达。后续可进一步研究这些基因的表达及相应次级代谢产物的变化,揭示黄酮类物质在波罗蜜CDM嫩茎抗病过程中的分子调控机制。
糖代谢在一定程度上能反映植株的生长状态[29-30]。谢柳青等[17]研究发现糖代谢对波罗蜜CDM茎次生生长有影响,且相比CK,CDM发生大量下调,少量上调。本研究通过KEGG功能注释及GO富集分析,也发现糖代谢相关基因大部分下调,且部分下调基因在光合作用途径较显著富集。据结果推测,糖代谢途径中关键酶的合成可能受到了抑制,而进一步影响茎的发育。后续可筛选出调节糖代谢的候选基因,鉴定与光合作用信号通路之间是否存在相互作用,以期为调控木本植物茎发育关键基因的研究提供参考。
蛋白质是生命活动的重要物质基础,翻译是执行其生物合成的重要步骤[31]。在非生物胁迫因子下,植物的生长发育受到不同的影响,还可能影响其基因的表达。大量研究发现环境胁迫能抑制正常蛋白质基因的表达,诱導逆境蛋白产生[32]。而付影等[16]研究发现突变体受环境胁迫程度较深。本研究通过KEGG分类及GO富集分析,结果发现27个和9个差异上调基因分别注释和显著富集在翻译途径,说明在波罗蜜CDM嫩茎中依然有蛋白质生成。推测在波罗蜜CDM嫩茎中产生了部分逆境蛋白,使其抵御逆境伤害。而关于波罗蜜CDM嫩茎的生长发育与逆境蛋白之间的调控关系,这是后续研究可继续探讨的问题。
参考文献
李志军, 段黄金, 吕春霞. 分枝列当茎的发育解剖学研究[J]. 塔里木农垦大学学报, 2000, 12(3): 12-14, 19.
Dharmawardhana P , Brunner A M , Strauss S H. Genome- wide transcriptome analysis of thetransition from primary to secondary stem development in Populus trichocarpa[J]. BMC Genomics, 2010, 11(1): 150-169.
Cosgrove D J. Growth of the plant cell wall[J]. Nature Reviews Molecular Cell Biology, 2005, 6(11): 850-861.
Sampedro J, Cosgrove D J. The expansin superfamily[J]. Genome Biology, 2005, 6(12): 242-252.
Liu J W, Hai G H, Wang C, et al. Comparative proteomic analysis of Populus trichocarpaearly stem from primary to secondary growth[J]. Journal of Proteomics, 2015, 126: 94-108.
Lucas W J, Groover A, Lichtenberger R, et al. The plant vascular system: evolution, development and functions[J]. Journal of Integrative Plant Biology, 2013, 55(4): 294-388.
Hohmann-Marriott M F, Blankenship R E. Evolution of photosynthesis[J]. Annual Review of Plant Biology, 2011, 62(3930): 515-548.
季道藩, 许馥华. 棉花叶绿素缺失的细胞质遗传[J]. 遗传, 1979, 1(5): 15-19.
刘振昌, 王莉青. 苦瓜白化苗生长发育调查分析[J]. 安徽农业科学, 2016, 44(34): 140-141.
Lu X M, Hu X J, Zhao Y Z, et al. Map-based cloning of zb7 encoding an IPP and DMAPP synthase in the MEP pathway of maize[J]. Molecular Plant, 2012, 5(5): 1100-1112.
Yamasato A. Tanaka R, Tanaka A, Loss of the N-terminal domain of chlorophyllide a oxygenase induces photodamage during greening of Arabidopsis seedlings[J]. BMC Plant Biology, 2008, 8(1): 64.
刘冬梅, 丁锦平, 李成伟, 等. 棉花突变体的获得及其应用研究进展[J]. 河南农业科学, 2009, 38(11): 11-15.
