β-伴大豆球蛋白α-亚基核心区的基因克隆、表达及纯化
2020-07-03李垚熹袁艳秋高梦楠周树义胡亚云栾广忠
李垚熹 袁艳秋 巨 倩 高梦楠 周树义 胡亚云 栾广忠
(西北农林科技大学食品学院,陕西 杨凌 712100)
大豆贮藏蛋白的主要成分为β-Conglycinin(7S大豆球蛋白),是一种由α′、α、β3种不同亚基构成的三聚体[1-2],占大豆蛋白的30%,其对大豆蛋白的结构功能特性影响显著。研究[3-5]表明,α′及α亚基的结构可分为核心区(core region)和延展区(extension region),且均含有两个N-连接糖基;β-亚基则不具备延展区,只有一个N-连接糖基。Maruyama等[6-7]研究发现,在N-连接糖基及延展区缺失后,原核宿主的核心区仍能自组装成三聚体。阻碍7S大豆球蛋白凝集的原因可能在于其糖基具有较强的亲水性[8-9],因此提高7S大豆球蛋白的凝胶能力的关键或许在于去除糖基。目前,重组蛋白纯化领域中起重要作用的是亲和标签,其中最普遍使用的是His标签。许妍妍等[10-11]采用基因克隆技术、基因重组技术以及蛋白分离纯化技术成功制备了缺失N-连接糖基的重组α’-亚基。试验拟选取品种为齐黄34的大豆种子提取大豆总RNA,使用大肠杆菌作为原核宿主对β-伴大豆球蛋白的α-亚基核心区进行克隆与表达,并利用AKTA蛋白纯化系统通过镍离子亲和层析柱对其进一步纯化,为后期重组大豆蛋白分散体系的构建提供依据。
1 材料与方法
1.1 原料、载体与菌种
大豆:齐黄34,山东省农科院;
感受态细胞E.coliDH 5α、E.coliBL21(DE3)、质粒pET-28a:西安热默尔生物科技有限公司;
质粒pGEM-T Easy:普洛麦格(北京)生物技术有限公司。
1.2 试剂与仪器
MiniBEST Plant RNA Extraction Kit、PrimeScriptTMII 1st Strand cDNA Synthesis Kit、DNA纯化试剂盒、质粒提取试剂盒、蛋白质分子量 Marker(3597Q)、rTaq DNA聚合酶、DL 2000 DNA Marker、DL 5000 DNA Marker、Quick Cut限制性内切酶EcoR Ⅰ、Xho Ⅰ:宝生物工程(大连)有限公司;
T4-DNA连接酶:普洛麦格(北京)生物技术有限公司;
琼脂粉、β-巯基乙醇、异丙基硫代-β-D-半乳糖苷(IPTG)、丙烯酰胺、十二烷基硫酸钠(SDS):北京索莱宝有限公司;
其余试剂为国产分析纯;
PCR扩增仪:Hema 9700型,珠海黑马医学仪器有限公司;
超微量紫外—可见分光光度计:NanoPro 2010型,北京鼎昊源科技有限公司;
冷冻离心机:Neofuge 15R型,上海力申科学仪器有限公司;
通风橱:CAV1000型,肯尼亚F&S Scientific公司;
超声波细胞破碎仪:VC130型,美国 Sonics & Materials公司;
蛋白纯化系统:ClearFirst-3000型,上海闪谱生物科技有限公司。
1.3 试验方法
1.3.1 引物的设计与合成 在NCBI(https://www.ncbi.nlm.nih.gov/)中查找7S大豆球蛋白α-亚基的基因序列,GB编号为NM_001249927.2(Glycine max beta-conglycinin alpha-subunit 〔CG-3〕, mRNA),依据编码核心区的基因序列设计一对特异性引物,即上游引物F1:5′-CCGACGACGACGACAAGGATTCTGAGTTACGAAGA-3′,下游引物R1:5′-CCGCTCGAGTTCAGTAAAAAGCCC-3′。考虑到蛋白纯化后N-端标签的处理,设计上游引物时引入了肠激酶酶切位点(加粗部分)及其保护碱基;为方便后续克隆,在下游引物中引入了内切酶Xho Ⅰ酶切位点(加粗部分)、终止密码子(斜体部分)及相应的保护碱基,理论上扩增所得的基因片段大小为1 300 bp左右,引物合成由宝生物工程(大连)有限公司完成。
1.3.2 目的基因的扩增 将齐黄34大豆种子使用液氮研磨成粉末,取300 mg冻干粉提取总RNA,而后以此为模板运用试剂盒自带的Random 6 mers构建cDNA单链。具体操作步骤:42 ℃逆转录1 h,70 ℃保温15 min,4 ℃恒温保存。以cDNA单链为模板利用自行设计的引物F1/R1对目的基因αc进行扩增,选用10 mg/mL的琼脂糖凝胶电泳对目的基因进行检测并回收,PCR反应体系参考Maruyama等[6]的方法修改如下:PrimeSTAR Max Premix(2×) 25 μL,F1(20 μmol/L) 0.5 μL,R1 (20 μmol/L) 0.5 μL,反转录反应液4 μL,无菌纯水补加至50 μL。PCR反应条件为98 ℃变性10 s,66 ℃退火5 s,72 ℃延伸30 s,35个循环,最后72 ℃延伸7 min。
1.3.3 重组克隆载体的构建、筛选及鉴定 参照许妍妍等[10-11]的方法并修改。通过T4-DNA连接酶将目的基因片段αc与选用的克隆载体pGEM-T Easy相连接,将连接体系于16 ℃反应1 h,导入感受态细胞E.coliDH 5α中。在含有氨苄青霉素的LB固体培养基中随机挑取4~5个独立的白色菌落用于菌落PCR筛选。菌落PCR反应体系:聚合酶rTaq mix 26 μL,单克隆菌株菌液2 μL,F1/M47 (20 μmol/L) 2 μL,F2/M48 (20 μmol/L) 2 μL,添加去离子水至50 μL。使用氨苄青霉素浓度为100 μg/mL的液体LB培养基对筛选出的阳性克隆菌株进行培养14 h,提取质粒,选用10 mg/mL的琼脂糖凝胶电泳及XhoⅠ/EcoRⅠ双酶切对所提质粒进行鉴定。首先通过XhoⅠ内切酶酶切重组质粒5 min,再加入EcoRⅠ内切酶继续酶切15 min。将鉴定结果与预期相符的质粒进行外送测序(由西安热默尔生物科技有限公司完成),将测序结果与NCBI的α-亚基核心区的基因编码区进行序列比对[12],将测序符合预期的构建成功的重组克隆载体命名为pGEM-αc,将阳性克隆菌株于37 ℃进行增菌培养后,于-80 ℃保存菌种。
1.3.4 重组表达载体的构建、筛选及鉴定 使用限制性内切酶XhoⅠ/EcoRⅠ对重组克隆载体pGEM-αc和选用的表达载体pET-28a进行双酶切,酶切反应体系:待酶切载体10 μL,内切酶XhoⅠ/EcoRⅠ各1 μL,10× Quick Cut Green Buffer缓冲液5 μL,添加去离子水至50 μL。将二者的双酶切体系于37 ℃反应15 min后回收产物,通过10 mg/mL的琼脂糖凝胶电泳对产物进行鉴定并回收具有双黏性末端的目的片段。
将上述两种目的片段回收后,在T4-DNA连接酶作用下于16 ℃连接1 h,连接体系(20 μL):pGEM-αc双酶切产物5 μL,pET-28a双酶切产物10 μL,50% PEG 2 μL,T4-DNA连接酶1 μL,T4-DNA连接酶10×Buffer 2 μL。将连接成功的产物导入到感受态细胞E.coliBL21(DE3)中,选用卡那霉素浓度为50 μg/mL的固体LB培养基对转化菌液进行培养,同时设置阳性与空白对照,37 ℃培养14 h。随机挑取若干独立的单克隆菌落,利用特异性上游引物F1和通用下游引物T7er对其进行菌落PCR筛选。将筛选出的阳性菌株接种于卡那霉素浓度为50 μg/mL的液体LB培养基中培养14 h,提取质粒,使用10 mg/mL琼脂糖凝胶电泳及Xho Ⅰ/EcoR Ⅰ单、双内切酶酶切对质粒进行鉴定,将鉴定结果与预期相符的质粒外送测序。以pET-28a-αc为测序结果正确的重组表达载体,以pET-28a-αc-BL21为阳性菌株,挑选单菌落的阳性菌株于37 ℃下进行增菌培养,菌种置于-80 ℃保存[13-14]。
1.3.5 重组蛋白的提取 将pET-28a-αc-BL21的菌液培养6 h,以体积分数为1%的接种量接种至含有卡那霉素50 μg/mL的液体LB培养基中,继续培养至菌液浓度OD600 nm值为0.8时,加入0.2 mmol/L的诱导剂IPTG进行诱导表达,于37 ℃持续培养9 h,取1 mL诱导菌液用于SDS-PAGE电泳检测(分离胶体积分数12%,浓缩胶体积分数5%),通过Quantity One软件分析目的蛋白表达量[15-17]。其余菌液于4 ℃,5 000 r/min离心15 min,收集菌体,使用磷酸盐缓冲液PBS (25 mmol/L磷酸钠缓冲液,500 mmol/L NaCl,1 mmol/L EDTA,1 mmol/L PMSF,0.02% NaN3,pH 7.4)对菌体重悬后通过冰浴超声破碎菌体[8],超声条件:振幅25,6 s/4 s,30 min。超声4~5个循环后取出,取1 mL全菌液用于后续SDS-PAGE分析,其余破碎菌体于4 ℃,8 500 r/min离心45 min,收集上清液即为目的蛋白α-亚基核心区的粗蛋白液,于4 ℃保存,将收集的全菌液、上清液和沉淀进行SDS-PAGE分析[18-19]。
1.3.6 重组蛋白α-亚基核心区的纯化 使用AKTA蛋白纯化系统对重组α-亚基核心区的粗蛋白液进行纯化,将制备好的100 mL粗蛋白溶液通过孔径为0.45 μm的水性滤膜过滤后,于4 ℃储藏,使用自动上样泵上样到预先用磷酸盐缓冲液平衡好的His Trap HP预装柱上,先使用5~10倍柱体积的磷酸盐缓冲液洗脱未与镍离子柱结合的杂蛋白,于280 nm下检测并收集各蛋白峰直至峰值达到基线,使用20%,50%,80%,100%的含有500 mmol/L咪唑的磷酸盐缓冲液对样品进行洗脱并收集对应的蛋白峰。将收集到的所有蛋白峰通过SDS-PAGE电泳进行分析鉴定[20-22]。
1.3.7 数据处理 使用NCBI官网的Blast (http://blast.ncbi.nlm.nih.gov/Blast.cgi)功能对测序结果进行对比分析;使用UniProt官网(http://www.uniprot.org/blast/)查找7S大豆球蛋白α-亚基核心区的氨基酸序列并使用Blast功能进行比对;使用DNA MAN软件分析重组表达载体的碱基序列及编码氨基酸;使用软件Snap Gene分析重组表达载体的测序结果;使用软件Quntity One分析目的蛋白α-亚基核心区的表达量;使用Image Lab分析核酸电泳凝胶图。
2 结果与分析
2.1 大豆总RNA的提取
由图1可知,1、3泳道的两个条带较为清晰,分别为28S和18S,证明提取的RNA并未被大量降解;而2、4泳道中的两个条带稍显模糊,尤其是2泳道中的条带弥散比较明显,说明所提取的RNA可能存在大部分被降解的情况,因此使用超微量紫外—可见光分光光度计进行进一步测定。1~4泳道的RNA样品浓度分别为96.65,35.22,85.94,53.95 ng/μL,各样品的A260 nm/A280 nm分别为2.04,1.75,1.97,1.95。综上,2、4泳道中提取的RNA样品质量不高,因此选取1、3泳道提取的大豆总RNA用于后续的RT-PCR试验。
2.2 目的基因的扩增
由图2可知,所得基因片段位置为1 000~1 500 bp,长度约为1 300 bp,与α-亚基核心区基因的理论值(1 265 bp)一致,证明α-亚基核心区基因片段扩增成功。

M. DL 5000 DNA Maker 1~4. 大豆总RNA

M. DL 5000 DNA Maker 1. α-亚基核心区基因片段
Figure 2 Amplification of target geneα-subunit core region by RT-PCR
2.3 重组克隆载体的构建、筛选及鉴定
由图3可知,采用自行设计的特异性引物F1/R1对随机选取的5个菌株扩增得到的片段长度均为1 300 bp左右,与目的基因的理论值相一致;而使用通用引物M47/M48扩增得到的片段长度为2 000 bp左右,是因为特异性引物与通用引物的位点中间有一定的片段大小,因此该结果也与理论值相符。理论上而言,重组克隆载体经单酶切后应形成清晰而单一的条带,但pGEM-αc经EcoRⅠ单酶切后形成两个清晰条带,其大小分别与克隆载体和目的基因的理论值相符,是因为选用的克隆载体pGEM-T Easy上带有EcoRⅠ酶切位点;重组克隆载体经Xho Ⅰ单酶切后能形成单一条带,证实重组克隆载体构建成功;而经双酶切后重组克隆载体被切成两个较为清晰的条带,且大小分别为3 000 bp与1 300 bp左右,与载体pGEM-T Easy(3 015 bp)和目的基因α-亚基核心区的理论值(1 265 bp)相符。
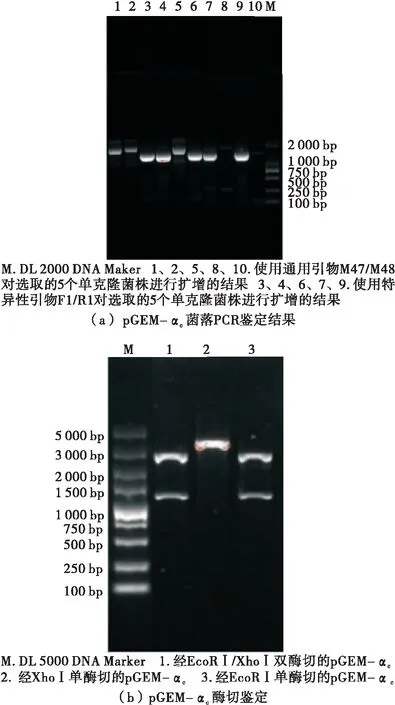
图3 重组克隆载体pGEM-αc的鉴定及筛选
Figure 3 Identification and screening of recombinant cloning vector pGEM-αc
2.4 重组表达载体的构建、筛选及鉴定
由图4可知,4、5、6、12泳道扩增所得产物条带大小为1 000~2 000 bp,约为1 300 bp,与目的基因理论长度相符,而其余泳道未出现目的条带,可能是因为挑选的菌落为假阳性。考虑到条带的清晰度、弥散程度和亮度,选用4、12泳道进行后续试验。pET-28a-αc被限制性核酸内切酶XhoⅠ、EcoRⅠ分别单酶切后均可得到与理论值大小相符的单一条带;而被限制性核酸内切酶XhoⅠ、EcoRⅠ双酶切后则得到两个条带,片段大小分别为1 300,5 000 bp左右,与α-亚基核心区(1 265 bp)和载体pET-28a(5 369 bp)的大小相符。通过测序比对可知,该序列与UniProt中编号为P0DO15的氨基酸序列相似度达到100%,未出现碱基移码和错配的现象,证明重组表达载体构建成功。
2.5 重组蛋白的存在形式
由图5可知,未添加诱导剂的工程菌pET-28a-αc-BL21经超声破碎后,其全菌液、上清液和沉淀中(1~3泳道)均未在50 kU附近出现目的蛋白条带;而37 ℃下诱导培养的菌体经超声后的全菌液、上清液和沉淀中(4~6泳道)均于50 kU处出现目的蛋白条带,由于上清液中所含的重组蛋白未形成包涵体,具有较好的溶解性,可收集用于进一步的分离纯化[13,23];蛋白纯化的穿透峰及洗脱峰浓缩后的滤液(8泳道)、诱导培养的菌体超声后的上清液经蛋白纯化的穿透峰及其浓缩液(9~10泳道)中无目的蛋白条带,与期望相符的,证明在使用低浓度咪唑洗脱杂蛋白时并没有目的蛋白被洗脱下来;而添加诱导剂的菌体超声后的上清液经蛋白纯化的洗脱峰及其浓缩液(11~14泳道)中有目的蛋白的单一条带,证明洗脱峰中目的蛋白含量较高,且目的蛋白含量为80%左右。
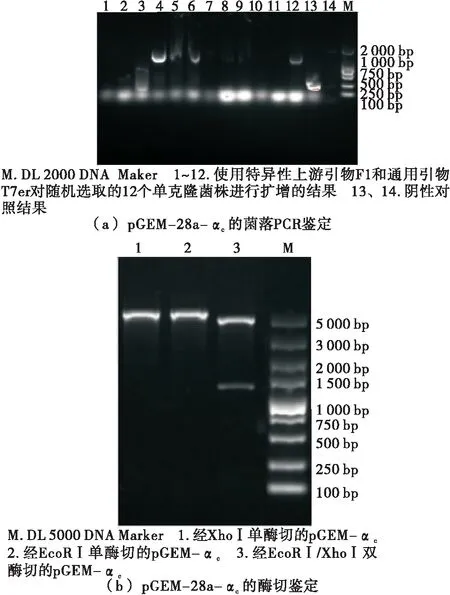
图4 重组表达载体pGEM-28a-αc的鉴定及筛选
Figure 4 Identification and screening of recombinant cloning vector pGEM-28a-αc
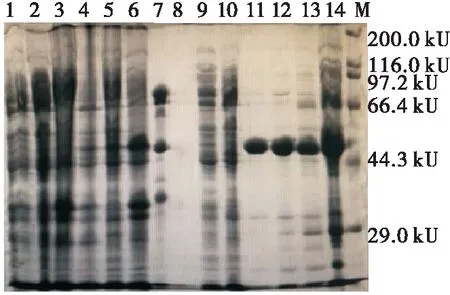
M. 蛋白质Marker 1~3. 未诱导工程菌的全菌、上清液和沉淀4~6. 诱导的全菌、上清液和沉淀 7. 天然7S 8. 蛋白纯化峰浓缩后的滤液 9~10. 蛋白纯化的穿透峰及浓缩液 11、13. 蛋白纯化洗脱峰 12、14. 蛋白纯化洗脱峰的浓缩液
图5 重组α-亚基核心区的提取及纯化电泳图
Figure 5 Electrophoresis of the extraction and purification of the recombinantα-subunit core region
3 结论
从齐黄34品种的大豆种子中提取总RNA,通过自行设计的特异性引物F1/R1经RT-PCR对7S大豆球蛋白α-亚基核心区的基因进行扩增,成功构建了经Xho Ⅰ/EcoR Ⅰ双酶切鉴定、菌落PCR鉴定以及碱基测序均正确重组克隆载体pGEM-αc和重组表达载体pET-28a-αc。将重组表达载体pET-28a-αc导入到感受态细胞E.coliBL21(DE3)中,经IPTG诱导表达获得分子量约为50 kU的重组α-亚基核心区蛋白,其表达条件为:OD600 nm值为0.8、IPTG浓度为0.2 mmol/L、诱导温度为37 ℃、诱导时间为9 h,此条件下重组α-亚基核心区蛋白可以得到大量表达,其纯度为87%以上。如何提高α-亚基核心区蛋白制备效率及纯度还需进一步研究。
