食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003对泡菜代谢物及风味的影响
2020-07-03徐亚洲谢书虎肖阳生刘长根
徐亚洲 熊 涛 谢书虎 肖阳生 刘长根
(南昌大学生命科学与食品工程学院食品科学与技术国家重点实验室,江西 南昌 330047)
泡菜发酵过程中的乳酸菌直接或者间接影响泡菜的风味代谢物质以及泡菜品质[1-4],已有研究[5]发现乳酸菌发酵泡菜卤水中风味代谢物与泡菜品质的差异,然而目前研究还未考虑到泡菜原材料携带的乳酸菌对泡菜品质的影响,缺乏乳酸菌泡菜发酵过程中组成风味物质的碳水化合物、有机酸、氨基酸以及挥发性物质动态变化的考察以及对风味成分的联合分析,尚未考虑基于感觉阈值差异的风味代谢物质对泡菜品质的贡献。试验拟对泡菜原材料灭菌处理后,以食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003作为起始发酵菌株纯种发酵泡菜,监测食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003纯种发酵泡菜及自然发酵泡菜中的代谢物的动态变化,通过风味物质的香气活度值与主成分分析考察不同发酵方式泡菜中代谢物与风味的特点,以期为泡菜品质改良与泡菜产业化发展提供理论依据。
1 材料与方法
1.1 材料、培养基与试剂
新鲜圆白菜、冰糖、碘盐、辣椒、姜、大蒜、花椒:市售;
MRS培养基:美国Sigma公司;
蔗糖、果糖、葡萄糖、乙酸、乙醇、乳酸:分析标准品,美国Sigma公司;
浓硫酸、无水乙醇、氢氧化钠、氯化铁、氯化钠:分析纯,上海国药集团化学试剂公司。
1.2 仪器与设备
pH计:pHS-25型,上海精密科学仪器有限公司;
液相色谱仪:Agilent 1260型,美国安捷伦公司;
气质联用仪:Agilent7820A气相-5977B型,美国安捷伦公司;
氨基酸分析仪:S-433D型,北京捷盛依科科技发展有限公司;
生化培养箱:DNP-9272型,上海精宏实验设备有限公司。
1.3 方法
1.3.1 乳酸菌培养与发酵菌剂的制备 从传统四川发酵泡菜中分离得到食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003,转接于MRS液体培养基中,37 ℃培养48 h后离心(5 000 r/min,10 min,4 ℃),用0.85% NaCl溶液洗涤两次,最后使用0.85%的NaCl溶液调节菌体浓度至9 lg (CFU/mL)。
1.3.2 泡菜的制备与取样
(1) 泡菜配方[6]:每100 mL水中,添加白菜50 g,冰糖4 g,碘盐4 g,辣椒4 g,大蒜3 g,生姜2 g,花椒1.5 g。
(2) 样品制备:泡菜原料混合均匀后分为4组,分别为食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003纯种发酵组,自然对照组与空白组,每个批次3个重复,自然发酵组不进行灭菌处理,室温下自然发酵,食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003纯种发酵组灭菌(105 ℃,25 min)后接入食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003纯种发酵,空白组原材料灭菌后不接入其他菌株,室温放置7 d。
(3) 取样:泡菜从入坛开始发酵7 d,期间每隔24 h于超净工作台内取样。
1.3.3 乳酸菌计数 取1 mL泡菜液加入到9 mL无菌生理盐水中,适当稀释后选择3个合适的梯度稀释液,分别取100 μL涂布于MRS固体培养基,每个梯度做3次平行涂布,37 ℃培养48 h。
1.3.4 pH值测定 采用pH计。
1.3.5 总酸含量测定 按GB/T 12456—2008《食品中总酸的测定》执行,总酸含量以乳酸计。
1.3.6 糖和有机酸含量测定 泡菜液于4 ℃,12 000 r/min离心10 min,取上清液,经0.22 μm滤膜过滤,滤液用于高效液相色谱分析。色谱条件:进样量20 μL,流动相5 mmol/L硫酸溶液,流速0.5 mL/min,柱温35 ℃,紫外检测器检测波长210 nm。选用示差折光检测器检测糖(蔗糖、葡萄糖、果糖)含量,紫外检测器检测有机酸(柠檬酸、乳酸、乙酸、琥珀酸、酒石酸、苹果酸、丁酸)含量。
1.3.7 游离氨基酸含量测定 取2 mL泡菜液,离心(3 000 r/min,5 min,4 ℃),取上清液1 mL,用2%磺基水杨酸稀释5倍,漩涡震荡30 s,静置15 min;离心(10 000 r/min,10 min,4 ℃);上清液用0.22 μm滤膜过滤,滤液用于氨基酸分析仪分析。
1.3.8 挥发性物质检测 泡菜中挥发性成分由GC-MS分离鉴定,色谱柱为HP-5MS毛细管柱(30 m×0.25 mm,膜厚度0.25 μm)。纯度99.999%的氦气作为载气,载气流速1.2 mL/min。烤箱温度起始为40 ℃保持1 min,以16 ℃/min的速度升至175 ℃并保持1 min,以2 ℃/min的速度升至95 ℃并保持1 min,以2 ℃/min的速度升至110 ℃并保持1 min,以3 ℃/min的速度升至150 ℃并保持1 min,以6 ℃/min的速度升至200 ℃并保持8 min。质谱离子源电压70 eV,离子源和传输线温度分别为230,250 ℃。质量扫描范围为m/z35~350。
1.3.9 代谢物香气活度值分析 泡菜中代谢成分决定着泡菜的感官风味,但是人类对风味代谢物质感官不仅取决于风味物质的相对含量,同时取决于风味物质的感觉阈值,而采用感官评定的方式误差较大,故依据泡菜风味代谢成分的香气活度值[7]评价不同发酵方式的泡菜风味。
(1)
式中:
O——香气活度值;
C——物质的绝对浓度;
T——感觉阈值。
由于Si=fi×Ci,所以
(2)
式中:
OA,OB——组分A、B的香气活度值;
f1,f2——气相色谱中组分A、B的峰面积的相对校正因子;
Si——组分i的峰面积;
Ti——组分i的感觉阈值。
风味物质的O值越高,则该风味组分对泡菜整体风味的贡献越大,结合主成分分析,对泡菜风味物质进行降维,乳酸菌纯种发酵泡菜过程中含量变化较大且不同发酵方式泡菜中差异较大的风味组分可作为泡菜风味的特征风味物质。
2 结果与分析
2.1 pH与酸度
如图1所示,总体来说,纯种发酵泡菜的pH值高于自然发酵泡菜,酸度低于自然发酵泡菜,可能是高温灭菌处理使原材料中的物质进入卤水中导致的[6],空白组中较高的pH值也从侧面证实了这一点。自然发酵泡菜的酸度大于纯种发酵泡菜,表明试验筛选的两株异型发酵乳酸菌产乳酸能力低于自然发酵泡菜中的多菌种发酵,Xiong等[8]对泡菜中乳酸菌的研究也得到了相似的结论。食窦魏斯氏菌NCU034005发酵泡菜的pH值高于柠檬明串珠菌NCU027003发酵泡菜,酸度小于柠檬明串珠菌NCU027003接种泡菜,说明食窦魏斯氏菌NCU034005发酵产乳酸能力小于柠檬明串珠菌NCU027003,该结果与图4(g)中显示的乳酸含量动态变化一致。自然发酵泡菜与柠檬明串珠菌NCU027003纯种发酵泡菜发酵后期盐水中的pH值变化趋于平缓,且pH值相差不大。柠檬明串珠菌NCU027003纯种发酵泡菜与食窦魏斯氏菌NCU034005纯种发酵泡菜的酸度相近,而自然发酵泡菜的酸度大于接种发酵泡菜,自然发酵泡菜是多菌种协同发酵,发酵后期,同型发酵菌株作为优势菌产乳酸[1],使自然发酵泡菜中的酸度较高,而食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003是异型发酵菌株,产酸与耐酸能力相对较弱,使泡菜发酵后期pH值趋于平缓[9]。柠檬明串珠菌NCU027003与食窦魏斯氏菌NCU034005对泡菜酸度的贡献较小,自然发酵泡菜中多菌种协同发酵有利于泡菜pH值迅速下降,致使泡菜迅速成熟,有研究[10-11]表明泡菜发酵前期以耐酸性较弱的球菌属为主,发酵中后期主要是耐酸性较强的同型发酵乳酸菌(如植物乳杆菌),与试验结果一致。
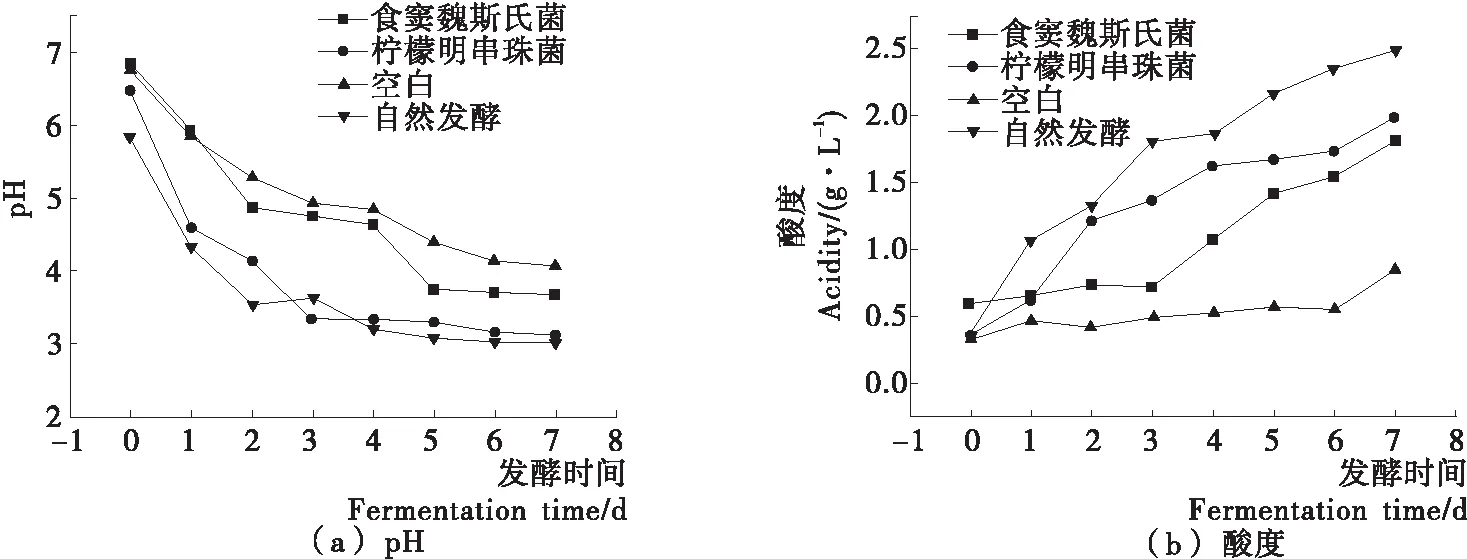
图1 发酵过程中泡菜pH与酸度的动态变化
2.2 乳酸菌纯种发酵泡菜过程中菌系的动态变化
如图2所示,泡菜发酵起始阶段纯种发酵泡菜卤水中乳酸菌数约为106~107CFU/mL,自然发酵活菌总数为103~104CFU/mL,而空白组卤水乳酸菌含量几乎为0 CFU/mL,且在整个发酵过程乳酸菌含量未有显著变化,说明灭菌效果良好,有效地排除了原材料表面附着的微生物对试验结果的干扰。乳酸菌纯种发酵泡菜中乳酸菌含量先上升后下降,可能是泡菜接种乳酸菌的作用,纯种发酵泡菜中食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003的乳酸菌数在早期迅速上升,有研究[9]表明异型发酵菌株耐酸性弱,导致了纯种发酵泡菜中乳酸菌数逐渐下降。自然发酵泡菜中后期乳酸菌含量高于纯种发酵泡菜,表明泡菜中耐酸能力较好的同型发酵乳酸菌逐渐增殖,与其他研究[12]结果相似,试验中筛选得到的食窦魏斯氏菌NCU034005耐酸性弱于柠檬明串珠菌NCU027003。
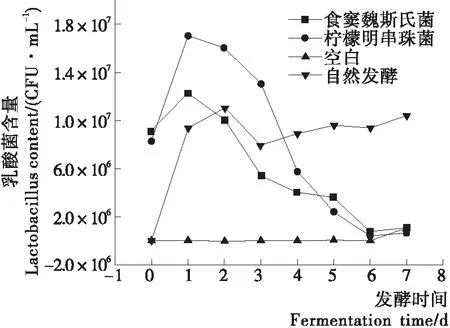
图2 发酵过程中泡菜乳酸菌活菌数的动态变化
Figure 2 Dynamic changes of viable lactic acid bacteria of paocai during the fermentation
2.3 乳酸菌纯种发酵泡菜过程中代谢物的动态变化
2.3.1 碳水化合物 由图3可知,蔗糖含量逐渐下降,果糖与葡萄糖含量逐渐上升,与Jeong等[13]的试验显示的游离糖含量均上升及Choi等[14]的试验显示所有的游离糖均下降有所不同,可能是试验所用的乳酸菌与原材料差异导致的。
如图3(a)所示,几种泡菜中蔗糖含量均在0~1 d中小幅上升,可能是泡菜原材料中的营养物质渗透进入卤液中导致的。自然发酵泡菜中的蔗糖含量低于纯种发酵泡菜,且食窦魏斯氏菌NCU034005纯种发酵泡菜中的蔗糖含量下降速度小于柠檬明串珠菌NCU027003发酵泡菜,说明自然发酵多菌种协同发酵对蔗糖的利用率高于食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003纯种发酵,柠檬明串珠菌NCU027003对蔗糖利用率高于食窦魏斯氏菌NCU034005,与其他研究[15-16]结果一致。
如图3(b)所示,空白样品中的葡萄糖含量基本不变,略有起伏,泡菜发酵过程中葡萄糖含量在0~1 d小幅下降,与图3(a)中蔗糖含量的动态变化一致,与其他研究[8,17]比较,泡菜中葡萄糖含量较低,可能由于泡菜发酵前期乳酸菌适应泡菜环境,乳酸菌活动消耗葡萄糖与原材料中蔗糖转化成葡萄糖的效率低于乳酸菌生成葡萄糖的效率。发酵前期,柠檬明串珠菌NCU027003纯种发酵泡菜与自然发酵泡菜中的葡萄糖含量逐渐上升,且柠檬明串珠菌NCU027003发酵泡菜中的葡萄糖上升速度大于自然发酵泡菜,食窦魏斯氏菌NCU034005纯种发酵泡菜中葡萄糖含量低于其他3种泡菜,由图2可知,纯种发酵泡菜中的柠檬明串珠菌NCU027003菌数在1~3 d高于自然发酵中乳酸菌数,食窦魏斯氏菌NCU034005转换蔗糖生成葡萄糖的能力远低于自然发酵,可能导致了不同发酵泡菜中葡萄糖含量的不同。泡菜发酵后期,4种泡菜中的葡萄糖含量动态变化趋于平缓,可能是泡菜中乳酸菌消耗葡萄糖的量与生成葡萄糖的量相当导致的。泡菜中球菌属乳酸菌一般可产生葡萄糖蔗糖酶将蔗糖转换成葡萄糖[9,18],加上泡菜酸环境以及乳酸菌自身活力的综合作用,导致了不同乳酸菌发酵泡菜中葡萄糖含量的差异。
如图3(c)所示,泡菜中的果糖含量动态变化趋势相似,发酵前期小幅下降,发酵中期逐渐上升,发酵末期趋于稳定。而自然发酵泡菜中果糖的含量总体低于纯种发酵泡菜,试验结果表明,食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003可代谢蔗糖生成果糖,转换效率相近。有研究[9]表明,球菌属特别是肠膜明串珠菌属是自然发酵过程中生成果糖的主要菌株。空白样品中果糖含量的变化可能是原材料中的果糖渗透进入泡菜液中导致的。
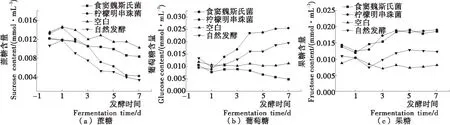
图3 发酵过程中泡菜蔗糖、葡萄糖与果糖含量的动态变化
不同异型发酵菌株代谢糖能力不同,冰糖的添加使泡菜中蔗糖含量增加,蔗糖作为冰糖中主要的糖类可以被蔗糖水解酶水解为果糖与葡萄糖,可能也是纯种发酵泡菜中糖含量不同的原因之一,乳球菌属尤其是柠檬明串珠菌NCU027003可分泌蔗糖转化酶将蔗糖转化成葡萄糖与果糖[9,19],且柠檬明串珠菌NCU027003可高效利用蔗糖与葡萄糖[20-21]。
2.3.2 有机酸 如图4所示,泡菜发酵过程中7种有机酸总体上升,柠檬明串珠菌NCU027003发酵泡菜液中除苹果酸外的有机酸含量都高于其他发酵泡菜。柠檬明串珠菌NCU027003发酵泡菜与食窦魏斯氏菌NCU034005发酵泡菜中丁酸、琥珀酸、酒石酸含量高于自然发酵,而自然发酵泡菜中柠檬酸、乳酸、乙酸的含量介于柠檬明串珠菌NCU027003发酵泡菜与食窦魏斯氏菌NCU034005发酵泡菜之间,发酵中后期苹果酸含量略高于柠檬明串珠菌NCU027003。发酵过程中酒石酸含量变化不大,可能是由于酒石酸不参与乳酸菌的生长代谢。柠檬酸含量在发酵中后期增长趋势趋于平缓,Koduru等[22-23]发现柠檬酸和葡萄糖与果糖、乳糖或木糖通过乙酸激酶途径代谢,泡菜中乳酸菌代谢柠檬酸,泡菜发酵中后期,柠檬明串珠菌NCU027003纯种发酵泡菜与食窦魏斯氏菌NCU034005发酵泡菜中的乳酸菌数降低,导致柠檬酸增长速度逐渐减慢。泡菜发酵过程中乳酸菌可以生成苹果酸,一些乳酸菌可以通过苹果酸乳酸酶将L-苹果酸脱羧为L-乳酸和CO2[24],导致了不同乳酸菌发酵泡菜中的苹果酸含量不同。
由图4(f)可知,柠檬明串珠菌NCU027003代谢产乳酸能力高于食窦魏斯氏菌NCU034005。泡菜发酵前期,接种乳酸菌使得纯种发酵泡菜中乳酸菌含量较高,导致了纯种发酵泡菜中乳酸迅速累积。研究[25]表明,同型发酵乳酸菌产乳酸能力高于异型发酵乳酸菌,自然发酵是多菌种协同发酵,发酵后期自然发酵泡菜中同型发酵菌是优势菌株,使得自然发酵泡菜中的乳酸含量逐渐接近柠檬明串珠菌NCU027003纯种发酵泡菜。
图4(g)中,柠檬明串珠菌NCU027003泡菜发酵过程中乙酸含量迅速增长,且高于自然发酵,可能是由异型发酵乳酸菌产乙酸较多,自然发酵泡菜中同型发酵乳酸菌产生乙酸含量较少导致的。有研究[25]表明,同型发酵乳酸菌代谢主要产乳酸,而异型发酵主要产乙醇、二氧化碳等物质。食窦魏斯氏菌NCU034005发酵泡菜中乙酸含量最少但高于空白样,结合图2可知,食窦魏斯氏菌NCU034005泡菜中的乳酸菌含量较低,故可能食窦魏斯氏菌NCU034005生成乙酸能力相对较弱导致的结果。总体来说,泡菜发酵过程中有机酸含量动态变化依赖于乳酸菌的活动,且乳酸菌发酵过程中乳酸含量高于乙酸含量,乳酸是造成泡菜酸环境的重要因素,与其他研究[25]结果一致。
2.3.3 氨基酸 如图5所示,泡菜发酵过程中谷氨酸(Glu)与天冬氨酸(Asp)含量逐渐上升,丝氨酸(Ser)与丙氨酸(Ala)逐渐减少。脯氨酸(Pro)、丝氨酸(Ser)、天冬氨酸(Asp)是发酵泡菜中含量较高的氨基酸,丙氨酸(Ala)在自然发酵泡菜中含量较高,其他种类游离氨基酸趋于稳定状态,总体变化不显著,而其他研究[13]发现发酵过程中氨基酸含量迅速增加,可能是由泡菜原材料、制作工艺、发酵菌株等因素导致的。
2.3.4 挥发性成分 如图6所示,在不同乳酸菌发酵泡菜过程中共检测出153种挥发性成分,包含32种醇、17种烯、15种酮、13种酯、10种生物胺与芳香化合物等挥发性物质,其中10种生物胺成分含量相对较低,主要存在于空白样中,且在泡菜发酵后期生物胺含量逐渐为0。
不同发酵泡菜中的挥发性物质组成有显著差别,食窦魏斯氏菌NCU034005发酵泡菜共检测出93种挥发性成分,柠檬明串珠菌NCU027003发酵泡菜共检测出55种挥发性成分,自然发酵泡菜共检测出43种挥发性成分,空白原材料共检测出了71种挥发性成分,食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003可显著提高泡菜挥发性成分的种类。桉树醇、松油醇、乙酸是3种发酵泡菜中主要的共有挥发性成分,樟树醇是食窦魏斯氏菌NCU034005发酵泡菜中独有的含量较高的挥发性成分,Jeong等[26]发现作为泡菜中重要的硫化物在试验中也被检测到。不同乳酸菌发酵泡菜产生的风味物质不同,而泡菜中多样的乳酸菌与其各自的代谢产物丰富了泡菜风味[27]。
2.4 代谢物的主成分分析
为了比较两种乳酸菌纯种发酵泡菜的代谢物的差异,对不同发酵方式泡菜中的代谢产物进行主成分分析,主成分1的方差贡献率为33.95%,主成分2的方差贡献率为50.91%。如图7所示,主成分分析载荷得分图表明不同乳酸菌发酵泡菜的代谢物有良好的分离效果,分别聚集在3个不同区域,在乳酸菌发酵泡菜过程中,泡菜中代谢产物发生了显著变化,空白组和食窦魏斯氏菌NCU034005泡菜的代谢物在发酵末期相近,柠檬明串珠NCU027003纯种发酵泡菜中的代谢物与自然发酵泡菜代谢物距离相近。说明异型发酵菌株柠檬明串珠菌NCU027003比食窦魏斯氏菌NCU034005对泡菜发酵代谢风味贡献更大。由图7(b)可知,A81(龙脑)、A22(樟脑)、A57(α-异松油烯)、A204(4-乙酸松油烯酯)、A33(醋酸)、A31(左旋香芹酮)、A91(桉树醇)、A153(3,7,7-三甲基二环[4.1.0]庚-3-烯)与苹果酸、蔗糖等是区分几种发酵泡菜差异的标志代谢物质。Nsogning等[28]也发现泡菜发酵过程中产生的醋酸是泡菜呈酸味的重要原因,而Hong等[29]发现泡菜发酵过程中形成的硫的衍生物在试验中不是区分柠檬明串珠菌NCU027003纯种发酵泡菜与食窦魏斯氏菌NCU034005发酵泡菜的标志代谢物,可能是使用的菌株不同导致的。不同发酵泡菜代谢物差异显著,柠檬明串珠NCU027003与食窦魏斯氏菌NCU034005以及自然发酵中的多种乳酸菌决定了风味各异的泡菜,不同乳酸菌有望产生品质各异的发酵泡菜。
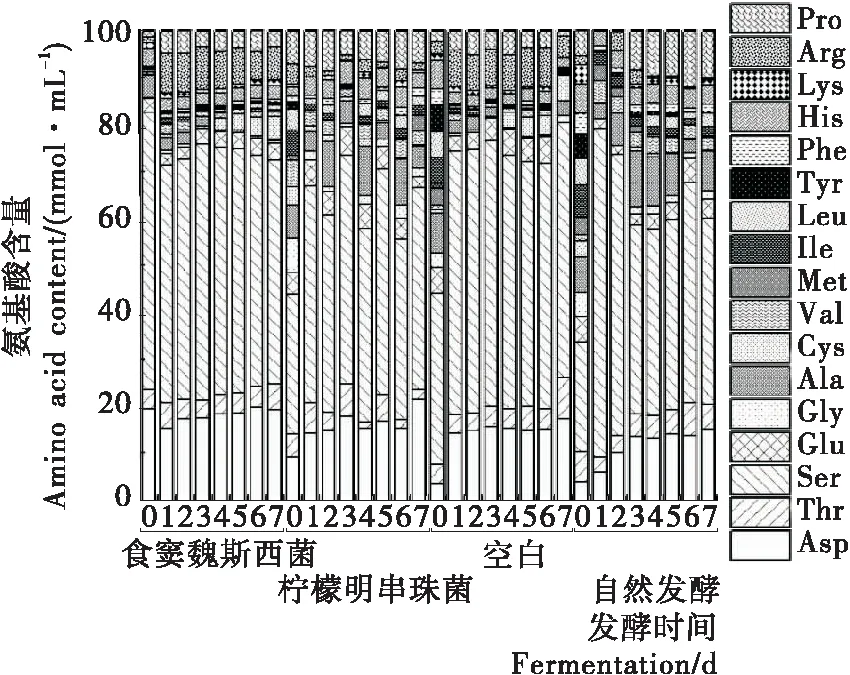
图5 发酵过程中泡菜氨基酸含量的动态变化
Figure 5 Dynamic changes of amino acid content during fermentation of paocai by different fermentation methods

图6 不同方式发酵泡菜中的挥发性成分
Figure 6 Volatile components of paocai fermented by different fermentation methods
2.5 泡菜风味的PCA分析
为了准确评估不同发酵方式泡菜的风味代谢物,找出乳酸菌纯种发酵泡菜过程中的特征风味物质,以自然发酵泡菜为标准,计算不同代谢物的香气活度值,评价食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003纯种发酵泡菜的风味,对乳酸菌纯种发酵泡菜中不同香气活度值的风味物质进行主成分分析,主成分1的方差贡献率为38.45%,主成分2的方差贡献率为48.94%。如图8所示,不同乳酸菌发酵泡菜风味物质显示了良好的分离效果,纯种发酵泡菜的风味与自然发酵泡菜有显著差异,食窦魏斯氏菌NCU034005发酵泡菜风味空白中风味相近,食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003对泡菜风味影响显著且具有不同的影响,A8(d-柠檬烯)、A1(α-松油醇)、精氨酸、醋酸与乳酸、酸度、果糖等风味物质是可显著区分泡菜风味差异的物质,不同发酵方式的泡菜中的酸度也有明显的区别,纯种发酵泡菜与自然发酵泡菜风味差异显著,这是由于自然发酵泡菜风味是多菌种共同作用的结果。泡菜中乳酸菌的活动产生了泡菜独特的风味,不同乳酸菌发酵泡菜的风味与品质差异明显[30-31],有研究[32]表明泡菜中风味的差别是由乳酸菌与酵母代谢产物引起的,还有研究[9]发现Leu.mesenteroides作为发酵剂可生产酸度更低、口味清爽的发酵泡菜,而试验中的食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003也可作为泡菜风味改良的菌种资源,但作为异型发酵乳酸菌产酸能力较弱,不能迅速使泡菜成熟。

图7 不同方式发酵泡菜代谢物的PCA分析
Figure 7 PCA analysis of metabolites of paocai fermented by different fermentation methods
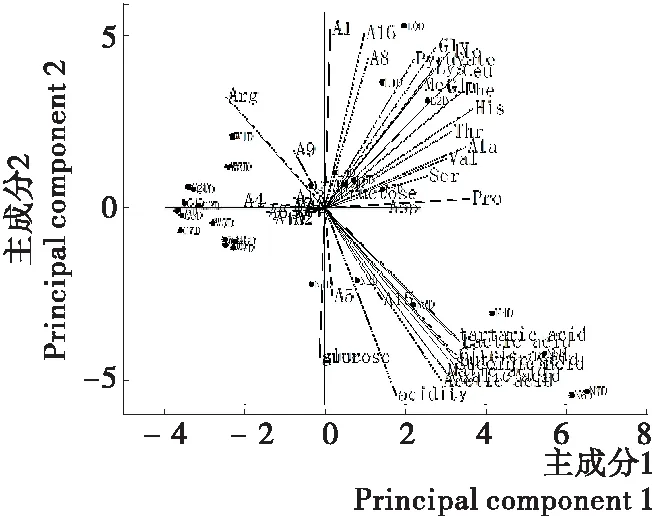
图8 不同方式发酵泡菜风味的PCA分析
Figure 8 PCA analysis of kimchi flavor fermented by different fermentation methods
3 结论
异型发酵乳酸菌食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003的耐酸性以及产酸能力较弱,两株乳酸菌对碳水化合物、有机酸与氨基酸的代谢能力不同,而食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003发酵泡菜各具有独特的风味,且与自然发酵泡菜风味差异显著,可认为是影响泡菜风味品质的重要菌种,其中龙脑、樟脑、α-异松油烯、4-乙酸松油烯酯、醋酸、左旋香芹酮、桉树醇等风味物质是泡菜中的特征风味因子,可作为监测泡菜品质的重要参考。食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003可作为泡菜风味改良的菌种资源。后续可进一步探究食窦魏斯氏菌NCU034005与柠檬明串珠菌NCU027003产特征风味物质的动力学关系以及其他种类乳酸菌纯种发酵泡菜的风味差异,探究乳酸菌发酵泡菜风味产生机理。
