精子阳离子通道1基因rs1893316位点单核苷酸多态性与特发性弱精子症的相关性▲
2019-11-21李伟伟闫娅妮殷秀荣
李伟伟 闫娅妮 刘 聪 马 波 殷秀荣
(河北省秦皇岛市妇幼保健院1 生殖医学科,2 优生遗传科,秦皇岛市 066004,电子邮箱:woshifeng-531@163.com)
世界卫生组织报告,全球育龄人群中不孕不育的发病率高达10%~15%,其中男方因素占不孕不育所有病因的40%~50%[1]。近年来,特发性弱精子症引起的男性不育已成为临床男性科的疑难杂症,其发病机制成为研究的难题。单核苷酸多态性(single nucleotide polymorphism,SNP)是最广泛的基因多态性,这些基因的不同可能会导致个体表型的差异、疾病的易感性等。目前,很多学者专注于男性不育与相关基因SNP相关性的研究,发现一些基因SNP可能参与细胞增殖和抗氧化过程,与精子发生过程相关,与男性不育存在一定的相关性[2-3]。精子阳离子通道(cation channel of sperm,CatSper)家族是精子特异性阳离子通道,共有4个成员(CatSper 1~4)[4]。CatSper1定位于鞭毛的主段,在增强精子运动、获能、超活化、受精等方面起重要作用,与弱精子症的发生相关[5-6]。本文旨在通过研究CatSper1的SNP与特发性弱精子症的相关性,进一步探索特发性弱精子症的发病机制,为治疗提供实验基础。
1 资料与方法
1.1 临床资料 纳入2016年5月至2018年6月在秦皇岛市妇幼保健院男科门诊就诊的192例特发性弱精子症患者作为弱精组。纳入标准:前向运动精子<32%;均正常性生活1年以上未育,其配偶无生殖相关疾病。另同期募集288例精液分析正常的已生育男性为对照组。所有研究对象均排除支原体、衣原体等感染,生殖激素和精浆生化检查均正常。弱精组年龄24~37(30.5±4.7)岁,体重(68.5±16.9)kg;对照组年龄25~36(29.8±6.1)岁,体重(71.3±23.8)kg。两组研究对象的年龄、体重差异无统计学意义(P>0.05),具有可比性。本研究经秦皇岛市妇幼保健院生殖伦理委员会讨论通过,所有研究对象均签署知情同意书。
1.2 精液的获取及常规分析 所有研究对象禁欲2~7 d,手淫法采集精液,收集于一次性无菌取精杯内,37℃水浴箱温育。待精液完全液化后按照世界卫生组织编写的《人类精液检查与处理实验室手册》第5版标准[7],利用计算机辅助精子分析(computer-aided sperm analysis,CASA)系统进行常规分析,并进行精子形态学分析。
1.3 精液CatSper1 rs1893316位点的SNP检测
1.3.1 引物设计与合成:登录GenBank网站(https://www.ncbi.nlm.nih.gov)查询CatSper1基因全序列AF407333,引物设计采用Primer 5.0软件完成,由上海索宝生物科技有限公司合成。引物序列如下:CatSper1上游引物为5′-ATTTACAATGAAGGAGTTTGACACA-3,下游引物为5′-GCCAGAGGAATAGTGGGATAAA-3′;内参甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)上游引物为5′-GGTATCGTGGAAGGACTC-3′,下游引物为5′-GTAGAGGCAGGGATGATG -3′。
1.3.2 DNA的提取:将500 μL精液与500 μL生理盐水均匀混合后,4℃下10 000 r/min离心5 min,洗涤沉淀后待用。采用核酸提取试剂盒(上海之江生物科技,批号:P20181201)提取DNA,在沉淀中加入100 μL核酸提取液,震荡混匀,置于100℃水浴中10 min。4℃下10 000 r/min离心10 min后洗涤沉淀,-20℃保存备用。
1.3.3 SNP的检测:将所有标本的DNA送至上海生物工程公司进行飞行时间质谱分析,采用Sequenom公司的MassArray生物芯片系统进行操作。(1)PCR扩增;(2)单碱基延伸;(3)树脂纯化;(4)芯片点样,启动MassARRAY Nanodispenser RS1000点样仪,将树脂纯化后的延伸产物移至384孔SpectroCHIP(Sequenom公司)芯片上;(5)质谱检测,将点样后的SpectroCHIP芯片使用基质辅助激光解吸附电离飞行时间质谱分析,检测结果使用TYPER 4.0软件(Sequenom公司)分型并输出结果。
1.4 精液CatSper1 mRNA的检测
1.4.1 标本处理:将液化后的精液缓慢加入密度梯度液上层,800 r/min离心15 min后洗涤沉淀,弃去上清,将沉淀混匀后轻轻加入装有0.5 mL培养液的试管底部置培养箱,将试管倾斜45°,静置30 min,收集上层云雾状精液用于mRNA及蛋白的检测。
1.4.2 mRNA的检测:利用Trizol试剂(Invitrogen公司,批号:10296010)提取总RNA。适量的焦碳酸二乙酯水溶解RNA沉淀,-70℃保存备用。取适量总RNA稀释后用分光光度计测定A260/A280,比值均在1.8~2.0,计算浓度后备用。使用反转录反应试剂盒(Fermantas公司,批号:K1622)进行反转录,操作严格按照说明书进行,产物于-70℃保存待用。建立25 μL PCR体系,包括2×Taq酶PCR反应混合液12.5 μL,20 μmol/L上下游引物各1.5 μL,模板DNA 2.0 μL,重蒸馏水7.5 μL。PCR反应条件为94℃预变性5 min,94℃变性30 s,56℃退火30 s,72℃延伸45 s,共35个循环,最后72℃延伸10 min,降至4℃保温。扩增片段长度为261 bp,凝胶电泳观察结果。使用Quantity-One软件进行分析,以目的基因与内参基因GAPDH灰度比值表示目的基因的mRNA相对表达量。
1.5 精液CatSper1蛋白的检测 将精液标本在室温下液化30 min,4℃下2 400 r/min离心15 min将精子与精浆分离,弃去上清液并用PBS洗涤3次。将沉淀重新悬浮,调节至含有50×106个精子/mL,置于1.5 mL离心管内,在4℃的CHAPS缓冲液{20 mmol/L Hepes(pH 7.4)、140 mmol/L氯化钠、10 mmol/L 3-[3-(胆酰胺丙基)二甲基]丙磺酸内盐、2 mmol/L乙二胺四乙酸、1 mmol/L乙二醇双四乙酸和完全蛋白酶抑制剂混合物}中匀浆,并在4℃下在热振荡器上孵育30 min。然后将裂解物进行15%十二烷基硫酸钠-聚丙烯酰胺凝胶电泳,并将蛋白质转移到硝酸纤维素膜(德国Roche Biosciences公司)上。将膜用5%脱脂奶粉在Tris缓冲盐水[10 mmol/L Tris-HCl(pH 7.2)及150 mmol/L氯化钠]中室温封闭4 h后与Tris缓冲盐水稀释的鼠抗Catsper1抗体(1 ∶500;Abcam公司,批号:ab165120)室温孵育过夜,用含有0.1%吐温20的Tris缓冲盐水洗涤3次后,再与辣根过氧化物酶标记的羊抗鼠的IgG(1 ∶4 000;Abcam公司,批号:ab150077)室温孵育1 h。显影仪快速显影后用Image软件对图像进行分析。
1.6 统计学分析 采用SPSS 25.0软件进行统计分析。采用拟合优度χ2检验对各组基因型进行Hardy-Weinberg平衡检验。计量资料以(x±s)表示,组间比较采用t检验;计数资料以例数或百分比表示,组间比较采用χ2检验;采用单因素Logistic回归模型进行危险因素分析。以P<0.05为差异有统计学意义。
2 结 果
2.1 两组精液常规分析结果比较 弱精组的前向运动精子比例低于对照组(P<0.05),而两组的精液浓度及体积、精子正常形态率差异均无统计学意义(均P>0.05),见表1。
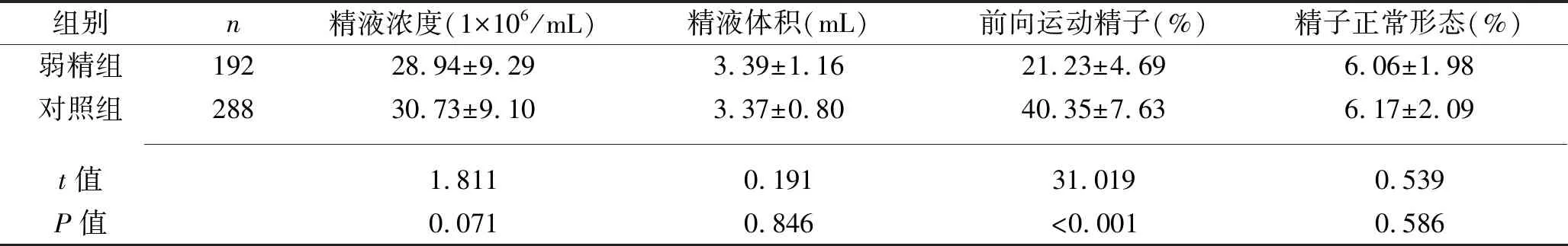
表1 两组精液常规分析结果比较(x±s)
2.2 两组CatSper1基因rs1893316位点基因型和基因频率、遗传模型比较 CatSper1基因rs1893316位点的基因型在对照组及弱精组的分布均符合Hardy-Weinberg平衡定律(均P>0.05),见表2。两组的CatSper1基因rs1893316位点基因型及等位基因的分布频率比较,差异均有统计学意义(均P<0.05),其中弱精组TT基因型、CT基因型及T等位基因高于对照组(P<0.05)。见表3。

表2 CatSper1基因rs1893316位点基因型的Hardy-Weinberg平衡检验结果[n(%)]
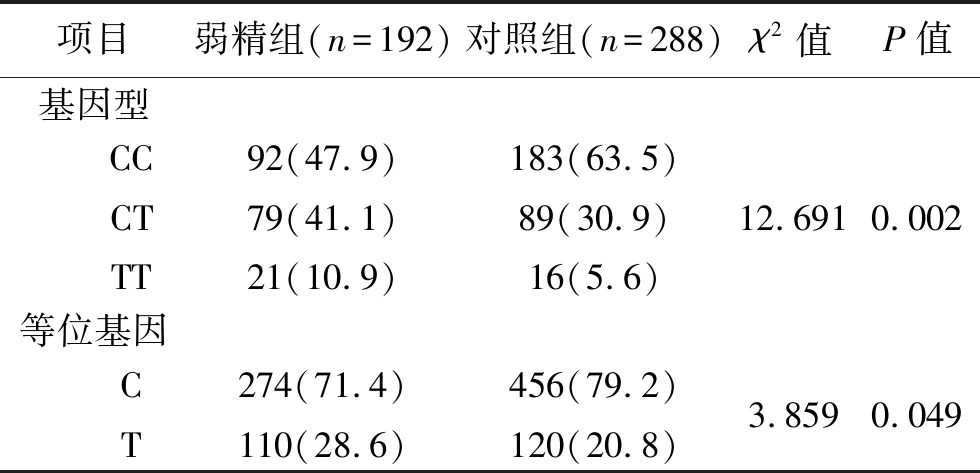
表3 两组CatSper1基因rs1893316位点基因型及等位基因分布的比较[n(%)]
2.3 CatSper1基因rs1893316位点多态性与特发性弱精子症的相关性 为了便于分析,将基因型多态性合并为二分类变量,产生遗传学显性模型、隐性模型和等位基因三种二分类结果。弱精组中,显性模型CC/(CT+TT)为92/100例,隐形模型TT/(CT+TT)为21/171例;对照组中,显性模型CC/(CT+TT)为183/105例,隐形模型TT/(CT+TT)为16/272例。以显性模型、隐性模型和等位基因为自变量(均设置哑变量),弱精症的发生作为因变量,进行单因素Logistic回归分析。结果显示,携带CatSper1基因rs1893316位点野生型CC是弱精症发病的保护因素,而携带突变型TT和等位基因T是弱精症发病的危险因素(均P<0.05)。见表4。
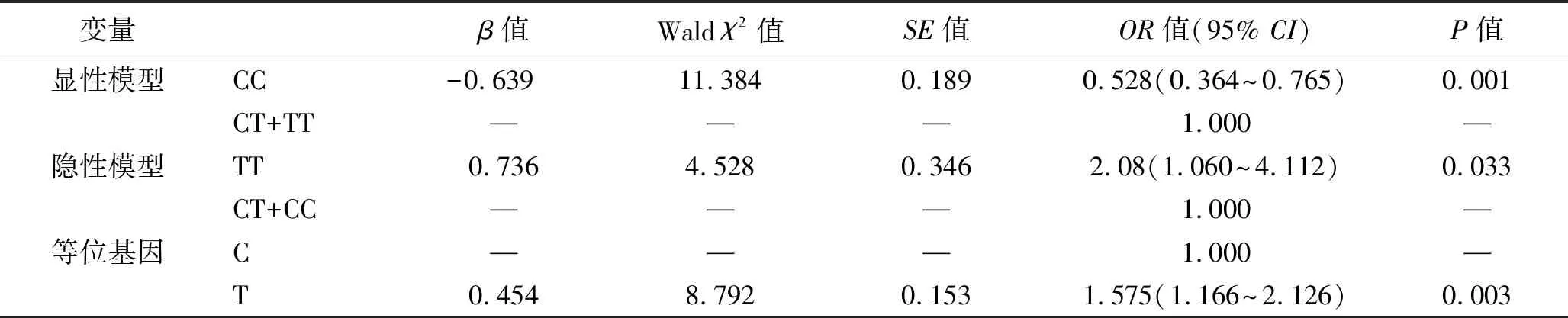
表4 CatSper1基因rs1893316位点多态性和弱精症发病风险的相关性分析
2.4 两组不同基因型携带者CatSper1 mRNA和蛋白的表达比较 携带TT基因型者中,弱精组的CatSper1 mRNA和蛋白相对表达水平均低于对照组(均P<0.05)。对于携带CC和CT基因型者,由于样本量很大,我们在对照组和弱精组中均随机各选取20例进行检测,结果显示CatSper1 mRNA和蛋白相对表达水平差异均无统计学意义(均P>0.05)。见表5及图1。
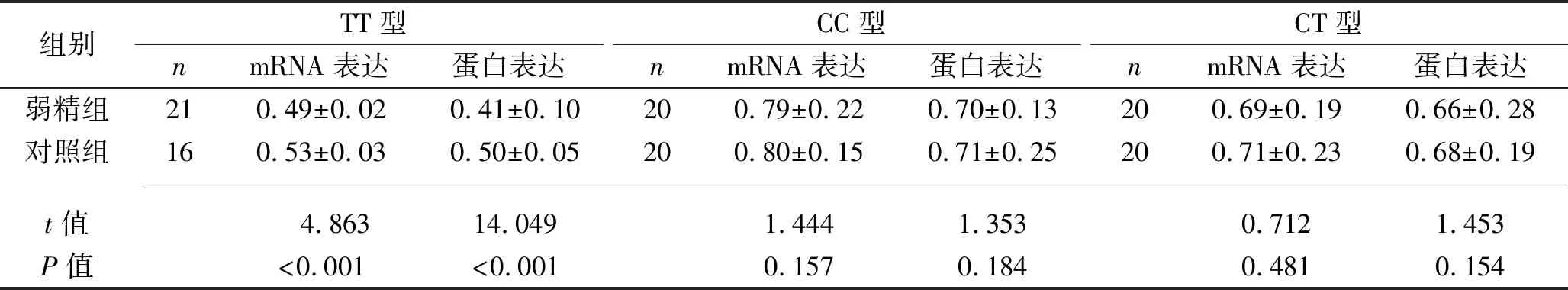
表5 两组不同基因型携带者CatSper1 mRNA和蛋白相对表达水平比较(x±s)
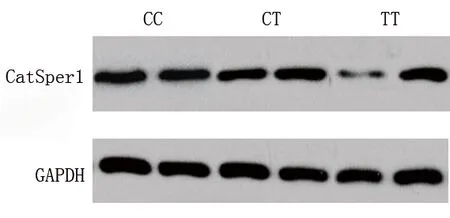
图1 携带不同基因型者精液CatSper1蛋白的表达情况
3 讨 论
近期有研究表明,精子鞭毛段和顶体主要含有一种特殊的蛋白质家族,称为CatSpers,其可以介导钙离子的流动,从而调节精子活力和过度活跃,在顶体反应和获能中起重要作用[8]。有学者发现。CatSper1敲除的小鼠精子活力较差,精子缺乏快速的前向线性运动以及环磷酸腺苷诱导的钙离子内流,并且鞭状运动减弱[9],提示Catsper1与精子活力密切相关。CatSper1敲除后精子不仅在获能后仍未见鞭状运动,也失去了穿透透明带的能力,精子尾部的钙离子内流消失[10],这表明Catsper不仅可以调节精子运动,还可以在精子过度活跃和顶体反应中发挥作用[11-12]。此外,临床研究表明,某些特发性不育症患者的精子活力下降与突变引起的Catsper1异常表达有关[13]。Wang等[14]研究发现CatSper1在附睾精子中下调或促进弱精子症的发生,而生精散治疗能够诱导弱精症模型大鼠的CatSper1上调和精子活力的提高。
近年来,已经有关于基因SNP与特发性弱精子症相关性的研究报道。例如,Buldreghini等[15]报告,编码内皮型一氧化氮合成酶中天冬氨酸(Glu298Asp)的T等位基因可能导致精子活力变弱。然而,很少研究关注CatSper 的SNP与特发性弱精子症之间的关系。虽然Visser等[16]报告了相关调查结果,他们只提供了序列数据,并没有对较大样本进行相关分析。因此,我们在Visser等的研究基础上,使用Sequenom的生物芯片系统,对192例特发性弱精子症和288例健康者精液CatSper1基因rs1893316位点的SNP进行检测。结果显示,弱精组TT基因型、CT基因型及T等位基因频率高于对照组(P<0.05);Logistic回归分析显示野生型CC是弱精症发病的保护因素,而突变型TT和等位基因T是特发性弱精症发病的危险因素(均P<0.05)。这提示CatSper1基因rs1893316位点的SNP与弱精子症的发生相关,可能是决定特发性弱精子症遗传易感性的重要因素,其中携带突变型TT和等位基因T者发生特发性弱精子症的风险更高。
作为同义SNP,rs1893316不改变CatSper1蛋白质结构。因此,我们推测rs1893316不通过影响氨基酸结构导致特发性弱精子症的发生。然而,位于CatSper1基因的第一个外显子中的rs1893316可能通过影响转录而导致特发性弱精子症。因此,我们评估了CatSper1基因rs1893316位点的SNP与CatSper1 mRNA和蛋白表达的关系,结果显示弱精组携带TT基因型患者的CatSper1 mRNA和蛋白表达水平均低于对照组携带TT基因型者(均P<0.05);然而,对照组和弱精组之间携带CC或CT基因型者的CatSper1 mRNA和蛋白的表达水平差异无统计学意义(均P>0.05)。这提示CatSper1 mRNA和蛋白表达下调,可能导致携带CatSper1基因rs1893316位点TT基因型者更易发生特发性弱精子症。
综上所述,CatSper1基因的rs1893316多态性位点与特发性弱精子症相关,携带TT基因型者发生特发性弱精子症的风险增加,这可能与CatSper1表达下调有关。
