猪链球菌2型安徽分离株多位点序列分型与毒力特征研究
2019-05-10张燎原汪清峰魏建忠
张燎原,陈 章,汪清峰,王 玮,孙 裴,魏建忠,李 郁
(安徽农业大学动物科技学院,安徽 合肥 230036)
猪链球菌(Streptococcus suis,SS)是导致猪链球菌病最主要的病原,该菌引起猪的败血症、脑膜炎、肺炎、多发性关节炎和多发性浆膜炎,人通过特定的传播途径也可以感染该菌。SS分成29个血清型,其中与疾病最相关的是SS2型(SS2),该型也是临床分离率最高的血清型[1-3],不仅给养猪业造成严重的经济损失,也给公共卫生带来威胁。SS毒力因子种类众多,其中胞外蛋白因子(Extracellular protein factor,EPF)、溶菌酶释放蛋白(Muramidase-released protein,MRP)、溶血素(Suilysin,SLY)是公认的3种主要毒力因子,不同地区分离菌株的毒力基因分布存在较大差异[4]。多位点序列分型(Multilocus sequence typing,MLST)是近年来用于细菌遗传进化分析的一种分子分型方法,具有很高的分辨能力和可操作性,可实现细菌流行病学跨区域监测。本研究对2009年~2017年分离的58株SS2安徽分离菌株进行epf、mrp、sly3种主要毒力基因检测、多位点序列分型(MLST)、斑马鱼和昆明鼠致病性试验,并综合菌株来源的临床病例分析,了解安徽地区SS2的毒力基因型分布和其ST型流行特点,以及毒力基因型、ST型与临床症状之间的相关性,为开展SS2流行病学研究,区分不同菌株间毒力差异以及促进疫苗的研发提供科学依据。
1 材料与方法
1.1 受试菌株来源 58株SS2安徽分离株源自2009年~2017年临床病猪,其中安庆2株、蚌埠11株、亳州7株、滁州1株、阜阳5株、合肥18株、淮北2株、淮南2株、六安6株、马鞍山1株、宿州1株、宣城2株,均由安徽农业大学动物传染病研究室分离、鉴定与保存。菌株名称以分离地区和时间命名。
1.2 主要实验材料及实验动物 2×TaqMasterMix、DL2000 DNA Marker、DL600 DNA Marker、细菌基因组DNA小量纯化试剂盒均购自TaKaRa公司;胰酪胨大豆酵母浸膏肉汤(TSB-YE)、小牛血清购自南京宝灵科生物技术有限公司。实验用约80日龄、体长约3.5 cm的AB系斑马鱼购自合肥裕丰花鸟市场御龙水族馆。5周龄18 g~22 g昆明鼠购自常州卡文斯实验动物有限公司。
1.3 引物合成 参照文献[5-6]合成SS的3种主要毒力基因epf、mrp、sly的引物;参照文献[7]合成SS的7个管家基因aroA、cpn60、dpr、gki、mutS、recA、thrA的引物。引物均由南京金斯瑞生物科技有限公司合成(表1)。
1.4 分离菌株3种毒力基因的检测 将冻干保存的52株SS2接种于含5%小牛血清TSB-YE液体培养基中,37℃培养18 h~24 h后提取其基因组DNA作为模板,用于PCR扩增 SS2的epf、mrp、sly3种毒力基因。反应条件为95℃5 min;94℃1min、退火温度(mrp:55℃、epf:60℃、sly:58℃)1 min、72℃ 2 min,40个循环;72℃ 10 min。PCR产物经琼脂糖凝胶电泳检测。

表1 基因名称及引物序列Table 1 Gene names and primer sequences
1.5 分离菌株MLST分型 根据文献[7]建立的MLST分型方法分别对7个管家基因进行PCR扩增,扩增产物由上海桑尼生物科技有限公司测序。将测序结果进行分析比对,获得菌株相应管家基因等位基因编号,确认相应ST型,并将新发现的等位基因序列提交MLST数据库。应用START v2.0软件构建UPGMA系统进化树作聚类分析,利用eBURST V3软件对各ST型进行亲缘关系分析并确定其克隆复合物。
1.6 分离菌株动物致病性试验 将受试菌株分别接种含5%小牛血清的TSB-YE液体培养基中,37℃培养14 h至对数期末到稳定期阶段,混悬液浓缩50倍后进行斑马鱼第一次感染试验,15尾/组,共52组。腹腔注射,10μL/尾,观察72 h后筛选出引起实验组鱼100%死亡的菌株;将该筛选出的菌株37℃再次培养14 h,分组进行斑马鱼二次感染试验,每组所用斑马鱼数量、注射剂量、观察时间等均与上同,筛选出引起实验组鱼100%死亡的菌株;采用平板计数法测定两次感染实验注射菌液的菌落总数。采用寇氏改良法分别测定筛选获得的菌株对斑马鱼、昆明鼠的LD50[8],比较受试菌株间毒力的大小,同时根据受试菌株与国内SS2强毒菌株HA9801、ZY05719对斑马鱼的LD50的比较结果[9],确定受试菌株的毒力。LD50测定试验中首先确定菌株对斑马鱼、昆明鼠模型的LD100和LD0,按照公式(n为分组数)计算公比,按公比设置各组剂量 ,分别为LD0、 LD0×r、 LD0×r2、 LD0×r3、LD100,斑马鱼实验组设置条件与感染试验相同,昆明鼠实验为10只/组,腹腔注射剂量为0.3 m L/只,一周内观察动物死亡情况。致病性试验全过程均设相应对照。
1.7 数据分析 使用X2检验,比较不同ST型在不同临床症状猪中的分布比例(肺炎、全身感染),p<0.05具有统计学意义。使用SPSS 22.0软件对相关数据进行统计学分析。
2 结果
2.1 分离菌株3种毒力基因检测结果 应用PCR方法检测58株SS2的epf、mrp、sly3种毒力基因。结果显示,分离菌株共分为5个毒力基因型:epf+/mrp+/sly+(26,44.83%)、epf+/mrp-/sly+(15,25.86%、epf-/mrp+/sly+(11,18.97%)、epf-/mrp-/sly+(3,5.17%)、epf-/mrp-/sly-(3,5.17%)(表 2)。表明epf+/mrp+/sly+、epf+/mrp-/sly+为安徽地区优势毒力基因型,在临床上更倾向于引起猪的全身感染(33/41,80.49%)。
2.2 分离菌株MLST分型结果 应用PCR扩增7个管家基因,上传数据库比对得到相应ST型,58株SS2安徽分离株包含10种ST型,分别为:ST1 26 株(44.83%)、ST7 18株(31.03%)、ST28 6 株(10.34%)、ST117 1 株(1.72%)、ST156 1株(1.72%、ST308 1 株(1.72%)、ST957 1 株(1.72%)、ST958 2株(3.45%)、ST959 1 株(1.72%)、ST970 1 株(1.72%)(表 2)。其中 ST957、ST958、ST959、ST970为新发现的ST型。表明安徽地区ST1、ST7为主要流行的ST型。依据菌株不同分离时间和地域分布进行统计,结果显示2009年~2012年江淮地区(合肥、淮南、滁州)分离菌株多数为ST1(7株)、ST7(3株)、ST957(1株)、ST958(1株);2013年~2017年以淮北地区(蚌埠、亳州、六安、阜阳、淮北、宿州)分离菌株最多,包括ST1 14株、ST7 6株、ST28 4株、ST117 1株、ST156 1株、ST308 1株、ST959 1株、ST970 1株(表3);9年间皖南地区(安庆、宣城、马鞍山)分离菌株数量较少,仅5株。包括ST1 2株、ST7 2株、ST28 1株。表明安徽地区SS2主要流行地域逐渐由江淮地区向淮北地区转变,自2013年以后ST分离数量明显增加,并伴随多种ST型出现。

表2 58株SS2安徽分离株毒力基因型与MLST分型结果Table 2 The virulence genetype and MLST identification of 58 SS2 Anhui isolates

表3 58株SS2安徽分离株分离时间和地域分布统计Table 3 Isolation time and geographical distribution statistics of 58 SS2 Anhui isolates
2.3 分离菌株ST型系统进化分析 应用START v2.0软件,基于7个管家基因串联序列构建UPGMA系统进化树。结果显示,58株SS2安徽分离株ST型可分为两个分支,分支 1由 ST1、ST7、ST156、ST308组成(46,79.31%),分支2由ST28、ST117、ST957、ST958、ST959、ST970 组 成(12,20.69%)(图 1)。4种新发现的 ST型 ST957、ST958、ST959、ST970 与 ST1、ST7、ST28、ST117、ST156、ST308之间亲缘关系较远。表明安徽地区SS2菌株流行的ST型遗传变化较为明显,且主要以分支1包含的ST型为主。
2.4 分离菌株ST型群落进化分析 应用eBURST V3将58株SS2安徽分离株的ST型与MLST数据库已有ST型一起进行聚类分析,7个管家基因同源较高的ST型归于一种克隆复合物(Clone complex,ST complex)。结果显示ST1、ST7、ST156属于ST1 complex,ST28、ST117 属于 ST27 complex,ST308属于 ST87 complex,新发现的 ST957划为 ST188 complex,其余ST958、ST959、ST970为单一ST型(Single)未被定义复合物归属(图2)。表明安徽地区ST1 complex、ST27 complex为SS2的主要流行复合物(86.20%,50/58),这与全球ST复合物流行趋势一致。

图1 58株SS2安徽分离株ST型等位基因序列的UPGMA系统进化树Fig.1 A UPGMA Phylogenetic tree based on ST allele sequence of 58 SS2 Anhui isolates
数据库中存在的ST型均用一个圆点(黄色)表示,绿色为复合物主要创建者,蓝色表明相应复合物名称,红色表示本实验包含的ST型。

图2 SS2全球MLST数据库eBURST聚类图Fig.2 Global MLST database eBURST clustering of SS2
2.5 ST型与感染动物临床症状的相关性 通过χ2检验,比较不同ST型在不同临床症状猪中的分布比例。临床症状以肺炎(分离自肺脏)和全身感染(分离自脑、脾、关节液部位)为分类依据。ST1、ST7、ST117、ST156和ST959菌株主要分离自全身感染病猪(37,80.43%),除ST117、ST959外均属于分支1包含的ST型 。ST28、ST308、ST957、ST958、ST970主要分离自肺炎感染猪(8,66.67%),除ST308外均属于分支2包含的ST型。在SS2可能引发的临床症状(肺炎、全身感染)中,分支1与分支2之间差异显著(p=0.0014),主要流行的ST1与ST7之间也呈显著性差异(p=0.0030)(表4)。表明分支1包含的ST型菌株更倾向导致以全身感染症状为代表的侵入性疾病,分支2包含的ST型菌株更倾向导致以肺炎症状为主要症状的呼吸道疾病;同时ST1较ST7更易于导致全身感染症状。
2.6 动物致病性试验结果 通过斑马鱼第一次感染试验,在58株SS2安徽分离株中筛选出10株引起斑马鱼100%死亡的菌株,再经斑马鱼第二次感染试验,进一步筛选出6株引起斑马鱼100%死亡的菌株,分别为ST1型2株(HF10-6、BB14-2)、ST7型4株(BZ13-4、HF13-8、XC09-2、HF09-6),且它们均属于ST1 complex。该6株菌对斑马鱼的LD50在0.62×102~3.39×104cfu之间,按照从小到大顺序依次为 : HF10-6<BZ13-4<HF13-8<BB14-2<XC09-2<HF09-6,均低于国内 SS2强毒菌株 HA9801、ZY05719对斑马鱼的LD50[9];对昆明鼠的 LD50在3.24×108cfu~3.72×109cfu之间,从小到大排序依次为:HF13-8<BZ13-4<HF10-6<HF09-6<XC09-2<BB14-2(表5)。结果表明,6株菌属于强毒菌株,毒力均强于国内强毒参考株HA9801、ZY05719,且HF10-6、BZ13-4、 HF13-8毒力强于BB14-2、 XC09-2、HF09-6。
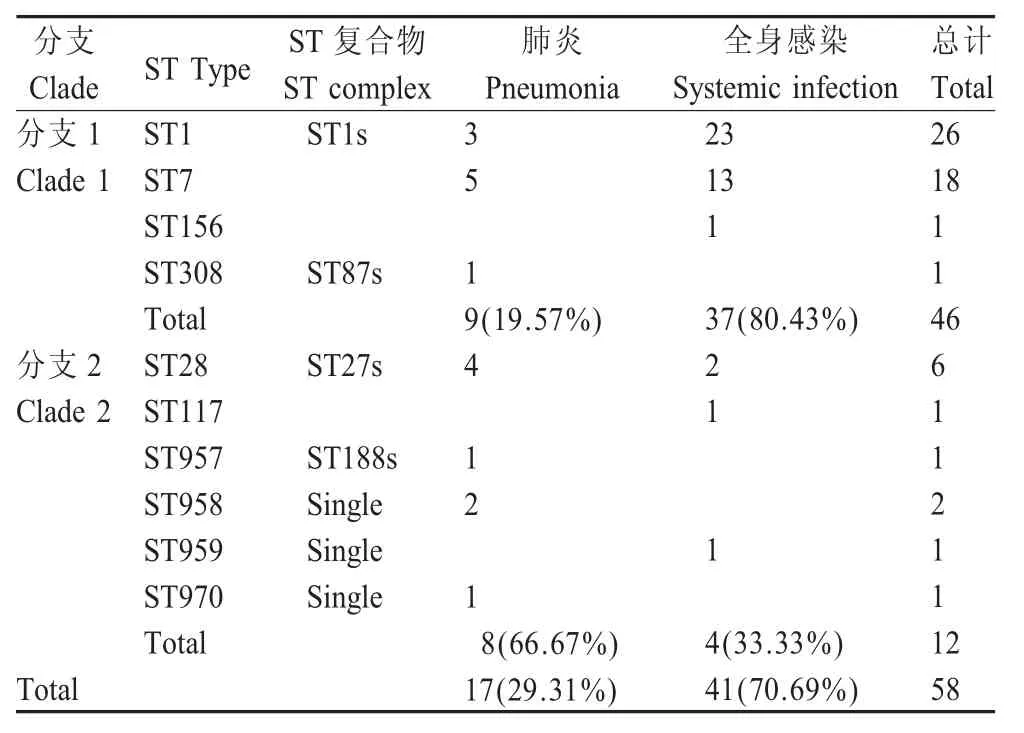
表4 58株SS2安徽分离株ST型与猪的临床症状关系分析Table 4 Association between STs and clinical characterization of 58 SS2 Anhui isolates
3 讨论
近年来分子分型技术的应用,极大的丰富了细菌流行病学遗传背景调查的手段,MLST作为一种具有高分辨率的分型方法,可实现跨区域性细菌流行病学监测,广泛应用于SS2的流行病学调查。Ye等通过MLST鉴定了2005年四川等地99株人源和猪源SS2菌株,其中98株为ST7型[10]。王楷宬等的研究结果显示,22株分离自四川、重庆、山东等地的SS2菌株均属于ST1型和ST7型[11]。Wang等对我国屠宰场生猪SS带菌情况的调查结果表明,ST1 complex和ST27 complex为主要流行的克隆复合物[12]。本实验针对58株SS2安徽分离株的MLST分型结果表明,在安徽地区主要流行的ST型为ST1、ST7(ST1 complex)和ST28(ST27 complex),与当前国内ST型流行情况一致。
从分离菌株来源的时间和地域来看,2009年~2012年安徽地区SS2主要流行ST1、ST7型;自2013年起首次出现ST28型,之后ST117、ST156、ST308型也陆续被分离到。在MLST数据库中查询ST型对应的最初登记信息,ST117为日本分离株DAT270,ST156为荷兰分离株GD-0036和德国分离株W 188.1,ST308则为国内分离株YS14、RC1。结果表明,在2009年~2017年的9年中,安徽地区SS2的ST型在不断增加,并伴有新的ST型产生,主要集中于淮北地区(包括蚌埠、亳州、六安、阜阳等地)。近年来,由于淮北地区生猪产能不断扩大,在生产交易和国际引种过程中,不同ST型的菌群间接触频率增加,可能存在不同菌群间等位基因的水平转移或随机的基因位点突变,导致ST型跨地域传播及新的ST型产生。通过聚类分析,新发现的ST957属于仅在中国区域存在的ST188 complex,表明区域内存在不同ST型间等位基因的水平转移,使得ST957与ST188 complex包含的ST型在7种管家基因中仅表现为1~2个基因的不同,并具有较高的同源性。
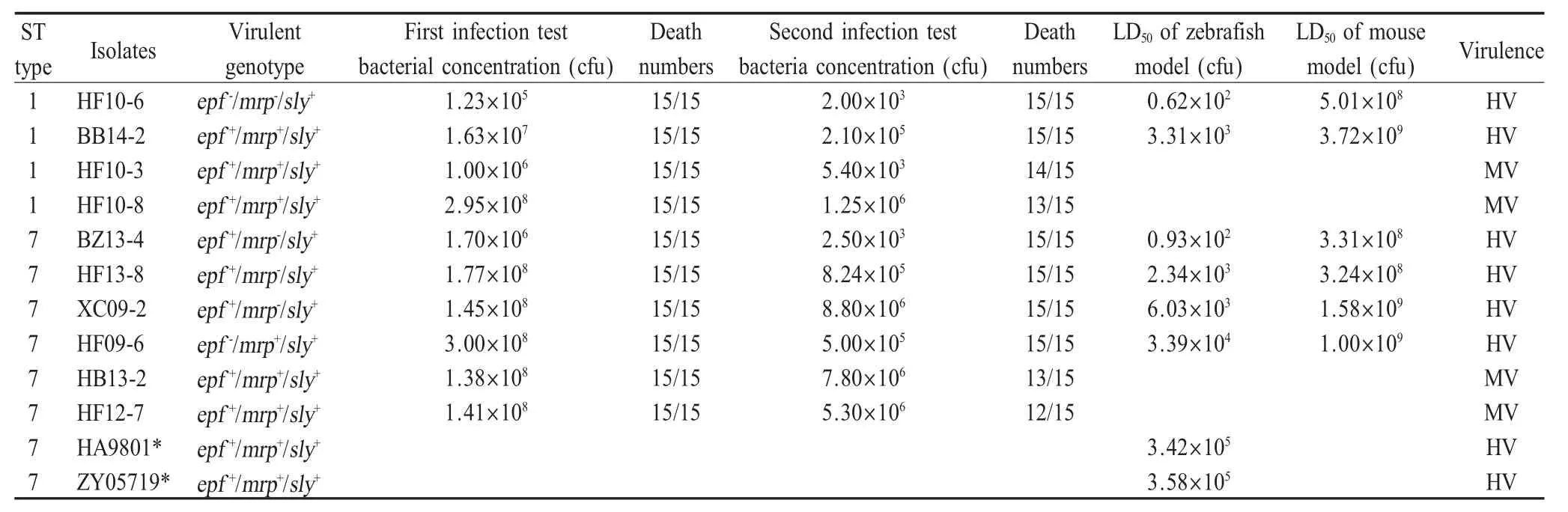
表5 10株SS2安徽分离株对动物感染试验结果Table 5 Artificial infection test of 10 SS2 Anhui isolates in zebrafish
Zhu等对180株中国SS2分离株进行MLST及毒力基因型分析,提出ST型与毒力基因型分布之间为随机性关联[13]。本实验也显示ST型与毒力基因型之间并无明确的相关性,但从流行趋势来看,ST1型/epf+mrp+sly+型仍是安徽地区SS2广泛流行的菌株特征。epf、mrp和sly作为SS 3种主要的毒力基因,虽然在分布上存在着全球区域性差异,但普遍认为epf+/mrp+/sly+型是我国SS2暴发流行的主要毒力基因型,并且与全身感染症状有密切关系[14]。结合临床症状,本实验显示安徽地区epf+/mrp+/sly+型SS2菌株倾向于侵入全身循环系统引起全身感染症状(92.30%),ST1 complex同样倾向于引起全身感染症状,主要包括脑膜炎、关节炎、败血症,并且与ST7型相比,ST1型这种倾向性更强,而ST27 complex则更易引起呼吸道疾病,该结果与相关文献报道一致[7]。本实验通过对斑马鱼、昆明鼠的感染试验和LD50测定,筛选出的6株强毒菌株均属于ST1 complex,表明安徽地区SS2的ST1 complex菌株中一部分菌株可能普遍具有强毒力特征。
分离菌株中ST1型菌株中的76.92%为epf+/mrp+/sly+型(20/26),ST7菌株中的55.56%为epf+/mrp-/sly+型(10/18)。临床导致的肺炎与全身感染的比例:ST1/epf+/mrp+/sly+型菌株为 2:16,ST7/epf+/mrp-/sly+型菌株为5:5。两种类型菌株主要差异在于是否携带mrp基因以及能否穿透血管壁进入血液循环系统导致全身感染。Li等研究显示SS2菌株中MRP可以与纤维蛋白原(Fg)相互作用增加血脑屏障(BBB)通透性,使菌体顺利穿过BBB[15]。表明MRP与全身感染症状密切相关,但上述统计结果显示并非所有mrp+菌株完全对应全身感染症状,应当存在其它毒力因子参与其中。King等报道在临床全身感染病例中,SS的sly+菌株分离率高于肺炎病例的分离率[16]。因此两类菌株携带的sly基因可能参与穿透血管机制。He等证实SS2菌株05ZYH33中SLY的高度表达促进其对上皮细胞的渗透是该菌株导致全身感染症状的主要原因,表明SLY也参与穿透血管内皮细胞[17]。此前研究已普遍认定SS菌株的致病作用是首先黏附和定居猪鼻、咽组织细胞,然后再扩散到呼吸道引发呼吸道系统症状,或进入血液引发全身感染性症状,虽然对细菌穿过血管的机制未有定论,但mrp、sly基因在其中的作用仍然为本实验中分析ST1/epf+/mrp+/sly+型与ST7/epf+/mrp-/sly+型在引起临床症状倾向的差异中提供参考依据。由于本实验分离菌株中的sly-菌株数量极少,未能有效论证sly基因在引起不同临床症状中起到的作用,所以仍需在今后的研究工作中作进一步探究。
本研究揭示了安徽地区SS2毒力基因型分布及ST型流行特点,结合动物致病性试验和菌株来源的临床病例信息,阐明了SS2毒力基因型、ST型与临床症状之间的潜在关联,探究了部分菌株在毒力、引起临床症状方面存在差异的可能因素。本研究为了解安徽地区SS2生物学特性、区分SS2菌株间毒力差异和流行病学研究提供了参考依据。
