运动力学刺激对骨质量影响的研究进展
2018-08-02赵常红李世昌孙朋刘媛胡晓磐
赵常红 李世昌 孙朋 刘媛 胡晓磐
1.西北民族大学体育学院,甘肃 兰州 7301241 2.华东师范大学“青少年健康评价与运动干预”教育部重点实验室,上海 200241
骨骼作为人体内最坚硬运动器官, 2017年发表在国际医学顶级期刊JAMA(美国医学学会杂志,影响因子44.405)的一篇文章指出,缺乏运动也是导致骨质疏松的主要原因[1],主要表现为骨质量下降,脆性增加,骨裂纹损伤非线性增高,骨重建功能下降,甚至出现微骨折,从而降低骨的生物力学性能。骨的质量包括骨量和骨结构两个方面,骨的力学性能也由这两个指标预测。骨量是决定骨力学性能的直接因素之一,骨量的减少是骨质疏松的一个重要标志。
运动力学的刺激使骨骼也不断调整、适应,运动力学刺激对骨髓间充质干细胞(mesenchymal stem cells,MSC)的增殖、分化、迁移,包括对骨微结构也有一定的影响。通过此综述来阐述运动生物力学影响骨形成质量和细胞形态的可能性机制。
1 运动力学刺激对MSC成骨分化的作用
MSC是一类具有自我更新和多向分化潜能的多功能干细胞,在损伤组织修复和再生中,MSC从骨髓中动员、进入外周血循环向损伤组织位点定向迁移是其行使损伤组织修复功能的关键环节之一。MSC的成骨分化有3个阶段:(1)快速增长阶段,形成胞外胶原基质;(2)基质成熟阶段,细胞增殖降低,碱性磷酸酶(alkaline phosphatase,ALP)活性增加;(3)矿化阶段,增殖进一步减慢,ALP活性降低,骨桥蛋白(osteopontin,OP)及骨钙蛋白(osteocalcin,OC)等成骨标志蛋白合成增加。成骨早期的标志基因ALP、成骨分化中期的标志基因OP和晚期标志基因OC可以结合羟基磷灰石,由成骨细胞(osteoblasts,OB)在基质矿化阶段合成[2]。
力学刺激是骨改建和生长的重要调节因素,在骨系细胞的分化与增殖中发挥关键作用。运动力学刺激在各种运动过程中完成各种动作给骨骼的一种反作用力。在没有力学刺激尤其是运动力学刺激对抗重力的情况下,会导致骨吸收增加,如卧床和失重。不同大小、频率和方式的力学刺激对骨组织的作用也是不同的,周期性的力学刺激比持续性的更能促进OB的增殖和基因表达,MSC在应力作用下可分化为OB[3]。研究发现,张力刺激可以促进MSC细胞增殖,提高其ALP的活性,显著上调成骨相关基因Ets-1、Runx2和ALP的表达,从而促使MSC向OB分化。研究还发现,MSC在力学刺激下,随着拉伸应变时间的增加,细胞的形态及伸展方向日趋沿着与力所施加的方向垂直排列,在骨质疏松症时MSC的成骨分化能力降低,成脂分化能力增高,导致骨髓中的脂肪组织增多,而运动力学刺激有助于降低成脂分化[4]。
MSC在受力刺激后形态、骨架的变化能调节其选择分化,以圆形形态生长时,MSC向脂肪细胞分化;当MSC以铺展和粘附形态生长时,向OB分化。已有实验证明,力学刺激细胞响应有个“阈值”,大于这个“阈值”,细胞才会应答反应,超过上限,就会有完全不同的、甚至负面的应答。较小的张应力能够诱导MSC向OB方向分化。有规律的力学刺激使得OB分化的标志物(Runx2、Osx、Col1)上升和脂肪分化标志物(PPARγ-2、C/EBPα和脂滴)下降,加强骨质疏松症患者的锻炼,已被证明是增加骨矿物质含量和阻止绝经后与年龄有关的骨矿物质、细胞外基质减少的有效方法[5]。应力介导下调MSC成脂或成骨分化,部分通过PI3K/Akt/GSK-3β信号通路[6]。研究发现,通过力学刺激可诱导MSC体外增殖,科学运动的力学刺激也可能调控体内MSC的增殖和去向分化,具有促进骨形成和抑制骨吸收的能力,能有效防治骨质疏松症且无药理方面不良影响。
2 运动力学对骨质量信号通路的影响机制
2.1 力学刺激对OPG/RANKL的作用
MSC的增殖与成骨分化成反比关系,细胞增殖快,分化就慢,反之,分化快,细胞增殖就慢。运动促进骨的重建是一个动态的过程,短期的力学刺激在短期内增加骨的形成。周期的重复性机械力的变化导致骨的微裂缝和维修重塑,引起一系列细胞反应,局部调整细胞因子、生长因子和其他分子,如:IL-1β、TNF-α、PGE2、IL-6、IL-8、RANKL、OPG、IGF、TGFβ-1和FGF[7]。此外,OB接受力学刺激表现出在分子水平如骨桥蛋白、骨钙素、PDGF和I、III型胶原的表达增加[8]。当运动生物力学刺激施加在OB时,它导致OPG/RANKL的表达升高,从而降低RANK/RANKL,抑制破骨细胞分化,包括p38 MAPK和ERK调节OPG感应介导转录因子负责RANKL表达的激活。破骨细胞受RANK调节,从而激活c-fos和NF-κB,这反过来又导致NFATc1活化。此外,运动形成的力刺激体外活性环氧合酶(cyclooxygenase,COX)和前列腺素的产生,减少RANKL从而抑制骨吸收[9]。力学刺激也被证明诱导的OB上的Wnt-β-catenin信号通路,促进OB的分化与骨形成[10]。见图1。
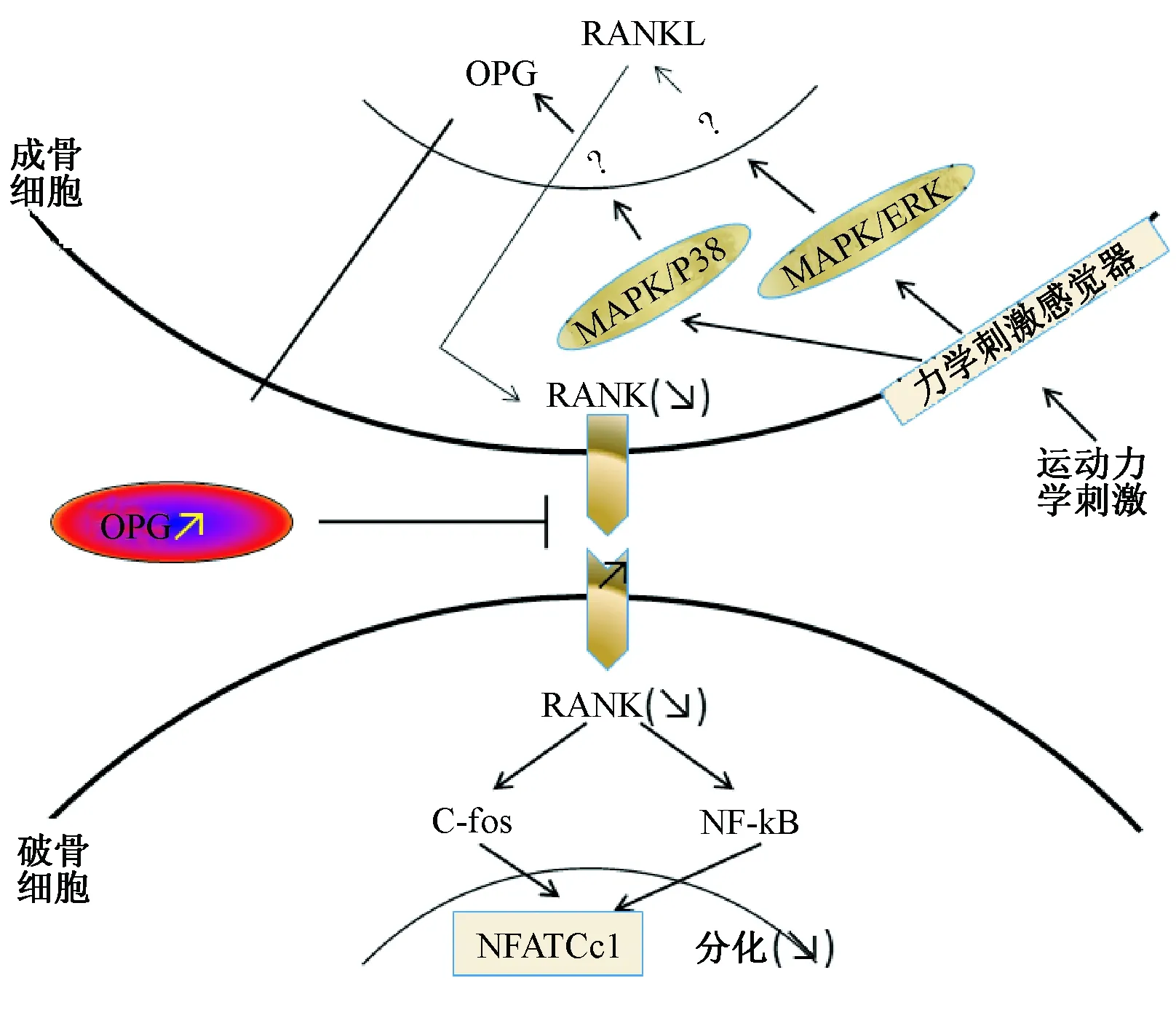
图1 在运动力学刺激下OB和破骨细胞相互作用的示意图[9]Fig.1 Schematic diagram of the interaction between osteoblasts and osteoclasts under mechanical stimulation of exercise[9]
2.2 力学刺激对MAPK/ERK的作用
研究发现,人OB样细胞在力学刺激后可直接促进脱氧核糖核酸的合成而不是通过自分泌,接收力学刺激的分子传感器在收到信号后传到细胞内部,开放的阳离子通道,OB间隙间交流及整合的产物也发现升高[11]。整合素是跨膜受体,将细胞外基质连接到细胞骨架,当一个信号出现时,与细胞骨架分子整合形成配合物。Rho家族成员的Ras相关G蛋白参与形成这些结构,除了上述作用,Rho家族成员可诱导多激酶和丝裂原活化蛋白激酶(mitogen activated protein kinase,MAPK)通路[12]。RhoGTP酶和其他相关Ras已被证明力在OB发挥作用,就力学信号而言,细胞外信号调节激酶(extracellular signal regulated kinase,ERK)和c-Jun氨基末端激酶(c-jun N-terminal kinase,JNK)似乎发挥至关重要的作用[13]。
ERKs已被证明在OB的生长和分化起重要作用,它们的激活对OB生存、增殖、分化有着不同的作用[14]。ERK和JNK信号通路已被证明会导致Fos和Jun增加,通过磷酸化激活它们,从而改变活化蛋白-1(activated protein-1,AP-1)的活性,增加Col1和OC的表达。力持续的时间、周期、强度是激活JNK/ERK基因表达的重要因素[15]。OB分化转录因子AP-1,调节Col1、骨钙素、OP和骨粘连蛋白的基因编码,这种转录因子在早期外部力学刺激反应中起重要作用。还有Fos和Jun家族成员,编码早期基因及MAPK信号通路的靶基因。OB可能在运动力学刺激的条件下,AP-1根据不同的细胞外信号调控基因转录[16]。研究表明,连续力学刺激人的成骨样细胞,激活MAPK级联反应,导致AP-1元件c-fos和c-Jun增加,更特别的是,连续应力导致成骨样细胞RABRho GTPases活性失调。另一方面,有研究发现,OB样细胞连续力学刺激后,AP-1作为一种重要的下游分子,通过p38 MAPK、MEK和RHoK通路调节表达,MAPK/ERK可能是一条重要的运动调控骨重塑和构建的通路[16]。
2.3 力学刺激对前列腺素E2(prostaglandin E2,PGE2)的作用
力学刺激会诱导成骨样细胞PGE2的产生,OB在生理应激、生长因子、激素、创伤和炎症细胞因子产生PGE2,进而诱导cAMP依赖的IGF-1产生[17]。IGF-1和IGF-2已被证明轮流诱发OBOSX的表达[18]。体内PGE2也会促使Runx2的表达升高,促进骨量增加,PGE2下游TGF-β在人成骨样细胞表达增加,在骨中具有增加细胞外基质的作用[19]。OB中包含TGF-β1的目标Runx2,这也是为什么Runx2基因敲除的小鼠成熟OB数量降低和细胞外基质的形成缺陷的原因[20]。因此,笔者也猜想运动的力学刺激是否可能通过诱导成骨样细胞PGE2的产生调控骨量的变化的呢?有待进一步研究。
2.4 力学刺激对一氧化氮(NO)的作用
OB对运动力学刺激的另一反应可能通过NO的产生,因为它结合到RAS调节位点,NO与MEK/ERK级联相互作用,会导致细胞增殖和细胞外基质的生产[21]。在后一阶段,Cox1、Cox2以及ERK1和ERK2被激活产生骨基质成分[22]。此外,力学刺激导致VEGF、BMP-2、BMP4、PGE2和IGF-1上调[23]。IGF-1通过PI3K-RACβ/Akt、MAPKs和Smad信号传导途径激活。在OB中BMPs诱导能促进Runx2、OSX和DLX5基因表达[24]。力学信号也能促进c-fos编码、早期生长反应因子1(Egr-1)和基本成纤维细胞生长因子(bFGF)基因的表达。力学刺激的性质和特性决定了骨的形成,体外骨细胞用高频和低强度的力学刺激,会导致细胞外基质增加,诱导骨形成,高强度力学加载促进BMPs拮抗剂表达从而抑制OB发育。连续力学刺激会导致细胞炎性因子及其受体的生产,如白介素(interleukin,IL)-1b,其生产与RANK-RANKL信号通路的激活相关,能促进骨吸收[25]。在生理强度水平内短周期的流体流动或循环的底物张力导致OB增殖和存活,能促进存活因子如IGF-1、IGF-2的产生和雌激素受体的激活,这也是重力维持OB活性的原因。失重或力学信号在体内不存在时会导致OB凋亡,减少与存活相关DNA转录因子的结合和正常线粒体失活,导致骨质疏松症发生,这也是运动防止骨质疏松的重要原因[26]。应力过度会导致细胞脱离表面导致体外程序性的失巢凋亡[27]。运动力学刺激促进骨的形成比较复杂,可能通过细胞的Ca2+通道、G蛋白、IGF、整合素受体、IGF和TGFβ/BMP相应信号通路的活化,从而诱导NF-κB、COX-2、CREB、Runx2、AP-1上调和通过Wnt抑制细胞成脂分化来提高骨的质量,如图2所示。
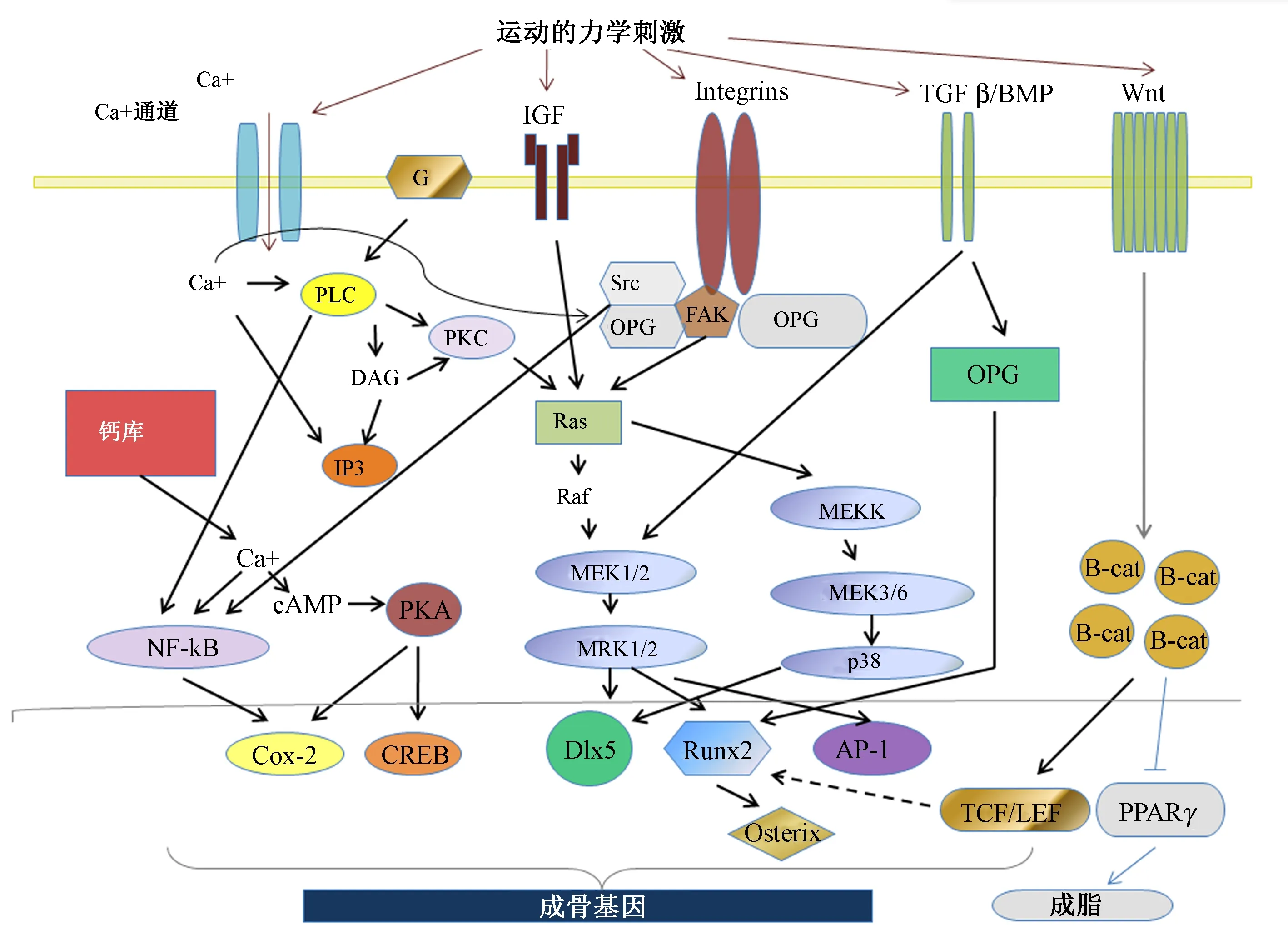
图2 运动力学刺激后的信号转导通路的交叉影响示意图[3,28]Fig.2 Schematic diagram of the cross effects of signal transduction pathways after mechanical stimulation of exercise[3,28]
2.5 力学刺激对多囊蛋白(polycystin,PC)的作用
最近研究发现,PC似乎与骨组织细胞的运动力学传导有关,如PC1,与原纤毛协同形成复合物,定位在特定的组织和细胞类型,包括OB、骨细胞。PC1基因敲除小鼠OB分化缺失,然后导致骨发育异常和骨质疏松,若PC1/原纤毛复合形成障碍,运动力学刺激的力学传导也可能会缺失。运动的力学刺激可能通过PC1调节细胞功能。通过JAK2/STAT3信号轴与PC1-CT互动,诱导人OB Runx2基因转录,调节成骨分化。运动力学刺激引发的PC1介导的磷酸化和STAT3的核转位,刺激诱导Runx2 mRNA升高。研究表明,力学刺激可能通过PC1-JAK2/STAT3信号轴上调Runx2基因表达,最终调控OB分化和骨形成[28]。
运动力学的传导研究虽然还不是很清楚,但原发性纤毛的协调PC已被认为作为潜在的OB力学分子感受器。研究表明,PC1的N-端可“接收”力学信号,通过其PC1的C端复合PC2共同定位在原纤毛,有接收力学信号的潜在功能。研究发现,通过激活G蛋白信号通路,PC1调节Ca2+,通过钙调神经磷酸酶/ NFAT轴传递信号,激活环孢素,一个特定的钙调神经磷酸酶/NFAT通路抑制剂,降低磷酸化NFATc1活性[29]。这些数据表明,PC1可能充当运动力学传导分子,通过钙调神经磷酸酶/NFAT信号级联调节OB的基因表达和OB分化。
2.6 力学刺激对Runx2的作用
Runx2转录因子在力学刺激向OB的分化和成熟中非常重要,是人成骨样细胞力学信号的目标基因。运动力学刺激可能直接导致Runx2表达和DNA结合能力的增加。运动力学信号从等离子膜也就是从整合素开始,穿过细胞质通过MAPK信号级联转导,靶向Runx2基因和蛋白。随着Runx2 DNA结合能力的增加,ERK1和ERK2被磷酸化,然后按时间顺序和平行的方式被激活。ERK1和ERK2磷酸化是相互的,在体内磷酸化和激活Runx2,依次诱导成骨分化[30]。
力学信号进入细胞可能通过整合和激活的Ras-ERK1/2 MAPK信号通路,导致Runx2磷酸化和转录活性增强。Runx2是通过自动调节机制控制自己的表达,运动力学刺激可能激活ERK的磷酸化和Runx2。这些分子结合到Runx2启动子促进Runx2的表达[30]。其次,Runx2基因启动子被发现附有典型AP-1结合位点,表明AP-1在调节Runx2的表达中发挥作用。此外,研究还发现AP-1和Runx2蛋白相互作用来调节胶原酶-3的表达[31]。
在失重或微重力情况下对骨造成的影响是毁灭性的。运动力学刺激对骨形成非常重要,能激活多种细胞内信号通路,其中激活的阳离子通道的流体剪切应力刺激ERK-CREB信号增强Fos家族转录因子的表达,增强Wnt/β-catenin信号,以此来调控Runx2上升。
2.7 力学刺激对NF-κΒ的作用
骨质量的变化不仅与骨的形成相关,也与骨的吸收相关,转录因子NF-κB与炎症过程有关,在破骨细胞的形成和骨吸收中起重要作用。NF-κB诱导骨重建和刺激破骨细胞的形成,它的激活通过RANK-RANKL系统或整合素将力学信号转换为Src激酶[32]。除上述功能外,NF-κB也可能在力学刺激后刺激OB分化。到目前为止,只有证据表明NF-κB在力学刺激后,OB在OB中的核中被激活和易位[33],运动力学刺激如何通过其激活OB特异性基因还有待进一步研究。
3 总结与展望
运动力学刺激可以有效增加骨矿物质含量,促进骨重建和塑造平衡,维持正常骨代谢功能。这一功能失调可导致骨质疏松症、甲状旁腺功能亢进症、甲状腺功能亢进症、佩吉特病,运动力学刺激维持这一功能在运动科学领域显得十分重要。虽然运动力学刺激分子机制已取得一定成果,但由于力学作用分子机制相当复杂,如:运动力的大小、持续时间、速度、频率及形式等的不同所产生的结果也会有所差别。因此,应不断关注力学刺激对骨质量影响在国内外的研究进展,为科学运动健骨提供理论参考。
