苏云金芽孢杆菌杀虫活性的增效机制研究进展
2018-02-01李超峰
李超峰
盐城师范学院, 江苏省滩涂生物资源与环境保护重点实验室, 江苏 盐城 224007
苏云金芽孢杆菌(Bacilliusthuringiensis,Bt)为革兰氏阳性昆虫病原细菌,可以产生具有杀虫活性的伴孢晶体蛋白,又称杀虫晶体蛋白(insecticide crystal proteins,ICPs)或δ-内毒素[1]。此外,α-外毒素、β-外毒素、γ-外毒素、热不稳定性外毒素和营养期杀虫蛋白(vegetative insecticidal protein,VIP)以及芽孢都有一定的杀虫活性[1]。然而,Bt毒素具有多样性,如根据Bt毒素晶体状态的不同可将其分为Cry毒素和Cyt毒素,而Bt又可分为不同的亚种,其毒素的种类和性质也不相同,从而使得其杀虫活性有较大差异[1]。鉴于人们对环境保护、食品安全和病虫害对化学农药抗药性认识的逐步深入,生物农药的研究和应用有了突飞猛进的发展。苏云金芽孢杆菌制剂以其高效、特异性强以及对环境、人畜安全等优势已经成为世界范围内应用最广、用量最大、效果最好的生物杀虫剂,因此,被广泛应用于农业、林业和贮藏等领域的害虫防治。
但在实际生产中,Bt制剂存在速效性差、持效期短、防效不稳定、杀虫谱较窄等诸多缺陷。为了克服这些缺陷,人们提出了一系列提高Bt毒性、增强防效稳定性的策略。如分离筛选杀虫活性高、特异性强的高效菌株;遗传改良现有高毒菌株;改进制剂制备技术;深度发掘各种高效生物、化学增效物质与增效因子等[2,3]。然而,大规模筛选高效菌株效率极其低下,且极为困难;而遗传改良菌株的稳定性差,在实践应用中难以实现。因此,快速、稳定、有效的增效物质、制剂和因子等的开发和应用受到越来越多的关注。只有清晰地了解增效物质、制剂和因子的作用方式和途径,才能更好地指导增效物质、制剂和因子在实践中的应用。本文在介绍Bt杀虫毒素蛋白作用机制的基础上,概述了不同增效方式与途径的研究进展,以期为苏云金芽孢杆菌生物制剂作用机制的研究提供一定的方向,同时,为增效剂在实际生产中的应用提供一定的理论基础和科学依据。
1 苏云金杆菌杀虫活性的作用机制
在田间,受长时间的高温、光照以及紫外辐射和电离辐射等的影响,大量羟自由基等活性氧的产生使细胞内生物大分子如DNA和蛋白质(包括Bt毒素蛋白)等的生物活性降低或者丧失。为了开发安全、高效的Bt生物制剂,明确其作用机制和增效机制尤为重要,因此,国内外对其展开了广泛而深入的研究。目前,被广泛接受的Bt作用模型有孔洞/顺序结合模式和信号转导模式[1,4~11]。
孔洞/顺序结合模式认为杀虫晶体蛋白被摄食后,昆虫肠道碱性环境将其溶解,进而被水解激活,55~65 kDa活化的Cry毒素与中肠上皮细胞刷状缘膜囊泡(brush border membrane vesicles,BBMVs)脂筏上的受体蛋白氨肽酶N(aminopeptidase N,APN)及碱性磷酸酯酶(alkaline phosphatase,ALP)结合、聚集,而后,再次与钙粘蛋白(cadherin)受体结合,使其构象改变,发生寡聚化,再与氨肽酶N 及碱性磷酸酯酶结合,进而插入上皮细胞,使细胞膜产生孔洞,裂解,最终导致昆虫死亡(图1)[1,4~6]。该模式需要多个受体的逐步参与,且能较好地解释杀虫蛋白与各受体结合能力的改变,或者受体表达水平的改变都能够导致昆虫对杀虫蛋白不敏感或敏感性降低的现象,即受体结合能力、表达水平或结构修饰均可以改变杀虫活性的敏感性。然而,在该模式中,如只表达多受体中的一种,也能够导致细胞对一种或多种毒素的敏感性降低[1]。
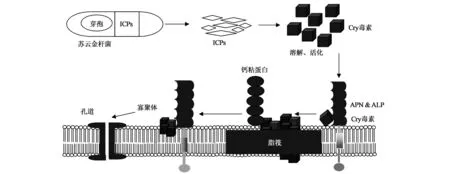
图1 Cry毒素作用的孔洞模式Fig.1 Cry toxin action model of pore formation.
信号转导模式完全不同于破坏细胞渗透压的孔洞模式,其认为活化的杀虫蛋白特异地结合了中肠上皮细胞上的受体钙粘蛋白,使得G蛋白(guanine nucleotide-binding regulatory protein)和腺苷酸环化酶(adenylyl cyclase,AC)被激活,磷酸腺苷环化可产生大量的环磷酸腺苷(cyclic adenosine monophosphate,cAMP),进而激活蛋白激酶A(proteinkinase A,PKA),引发细胞骨架重排、离子流动等一系列变化以及第二信号途径的加快,最终导致细胞死亡。同时,活化的杀虫蛋白与受体的结合,增加了钙粘蛋白从细胞内膜小囊泡到细胞膜的定向运输(图2)[7~10]。该模式依赖于Mg2+催化三磷酸腺苷(adenosine triphosphate,ATP)向cAMP转化,Cry1Ab杀虫蛋白可导致细胞形态改变,如膨胀、破裂,而二价阳离子螯合剂可以阻止Cry1Ab对表达钙粘蛋白的昆虫细胞的破坏,同时,细胞中也存在对杀虫蛋白信号转导的防御机制,可对损伤的部分上皮层表皮细胞进行修复[7,9,11]。因此,该模式忽略了其他系列受体的作用,也难以解释与受体结合能力未发生改变的杀虫蛋白突变体没有杀虫活性的现象。
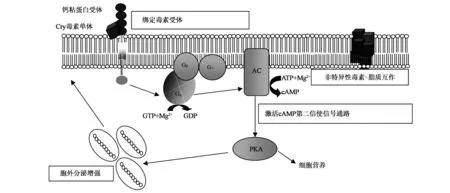
图2 Cry毒素作用的信号转导模式Fig.2 Cry toxin action model of signal transduction.
2 苏云金杆菌杀虫活性的增效方式
2.1 增加Bt制剂的摄入量,提高其有效利用率
无论何种寄主植物,绝大多数昆虫对Bt都具有一定的拒食作用,且根据次生代谢产物组成的不同,昆虫对寄主植物的取食嗜好也有所差异,对附着于植物器官表面的杀虫蛋白等毒素的摄食量也不同,昆虫摄食的杀虫蛋白等毒素不充分,进而导致杀虫效率降低[12,13]。棉籽粉、蔗糖、葡萄糖、粗棉籽油和吐温80等取食刺激剂和表面活性剂可通过增加目标昆虫的适口性,或降低生物制剂水溶液表面张力来改善药液在植物叶面的附着效果等,以提高Bt制剂的有效利用率,减少使用量,从而达到增效作用[14]。
2.2 改变目标昆虫中肠pH环境,提高原毒素溶解和活化的机率
摄入的原毒素只有经过溶解和活化才能启动杀虫活性,目标昆虫中肠的碱性、还原性环境,与蛋白酶的共同作用将实现其溶解和活化。因此,中肠环境和内容物对原毒素的溶解和活化是非常关键的,也是实现杀虫活性增效作用的第一个环节。化学添加剂中碱性化合物、蛋白质助溶剂和二价金属离子等分别可使敏感昆虫中肠的碱性提高(pH增大)、还原性增加,以及蛋白酶被激活和活性提高,从而使原毒素的溶解和活化得以促进,达到活性增效的目的[2,15]。其次,除了可使杀虫蛋白的溶解加速,无机盐、丝氨酸、精氨酸、丙氨酸、色氨酸、已酰胺、硝酸钠等多种含氮化合物还可以改变昆虫血液、淋巴液的酸碱环境和化学组成,影响正常的生理代谢过程,提高其对病原的敏感性,从而改善杀虫蛋白的毒效,进而达到增效目的[2,15~18]。此外,硼砂、硼酸、单宁酸和间苯二酚等具有适度毒性的化合物也可对Bt产生增效作用,但这些化合物对Bt生长具有一定的抑制作用,因此,作为复配剂时需要注意其组成比例[17,18]。
2.3 改变毒素蛋白的结合活性,增加毒素蛋白的利用率
原毒素被激活后,要经过与受体结合、插入上皮细胞,并形成孔洞或离子通道等一系列过程才能实现杀虫活性,因此,该环节任何一步的改变都将影响杀虫效率。首先,增效剂通过降低中肠蛋白酶水解活性,增加毒素蛋白与受体识别机率,促进具有良好插入能力的杂合寡聚体的形成与聚集,加速昆虫上皮细胞受体孔道的形成,离子渗透能力加强,从而实现对杀虫蛋白的增效作用[6,19~29]。其次,Cyt1Aa杀虫蛋白具有高度亲脂性,在体内结合到中肠微孔膜后,为Cry蛋白提供了额外的受体结合位点,提高了Cry毒素的结合机率,触发了插入膜囊前的成孔寡聚前体结构的形成,进而促进成孔、离子通道的形成和毒素作用的发挥[6,19,21,30]。苏云金芽孢杆菌可以产生不同的杀虫蛋白,不同杀虫蛋白的毒力和功能都不同,而不同功能的杀虫蛋白所产生的联合作用较单一毒素更为有效[5]。Tabashnik[31]提出了测定联合作用的方法并证明了Bt以色列亚种27 kDa CytA毒素和130 kDa或65 kDa的Cry4对消灭埃及伊蚊具有增效作用,而在2种Cry4毒素间或Cry1A不同毒素间无协同增效作用。Bt 4Q7菌株表达的135 kDa Cry1Aa和130 kDa Cry1A 2种毒素混合物对消灭甜菜夜蛾(Spodopteraexigua)和棉铃虫(Helicoverpaarmigera)都具有增效作用[26]。与大肠杆菌(Escherichiacoli)共表达的130 kDa Bt以色列亚种Cry4Ba和29 kDa Bt达姆斯塔特亚种 Cyt2Aa2对消灭埃及伊蚊和致倦库蚊(Culexquinquefasciatus)具有协同增效作用[22]。
δ-内毒素的不同毒素蛋白间的协同作用机理较为复杂。如Bt以色列亚种可产生不同的Cry和Cyt毒素,每种毒素对蚊幼虫都具有杀虫活性,而共同作用时则具有协同增效作用。这种协同增效作用是由于Cyt1Aa为Cry11Aa提供了额外的结合位点,使得插入细胞膜通道前体高效形成,进而促进离子通道的形成和与目标膜的结合,从而实现对Cry11Aa的增效作用,并降低害虫对Cry11Aa杀虫蛋白的抗性[6,21]。同样,Cyt1A98毒素通过为Cry4BLB毒素在埃及伊蚊(Aedesaegypti)BBMVs提供额外的结合位点,促使Cry4BLB杀虫蛋白寡聚结构的形成,从而实现其协同增效作用[23]。而Cry4Ba毒素则通过发夹结构β2-β3区域Thr328残基和β4-β5区域Thr328残基增强与Cyt2Aa额外受体的结合,实现其协同增效作用[20]。除了不同杀虫蛋白间的协同增效作用,中肠受体蛋白多肽如钙粘蛋白多肽和蛋白氨肽酶N(aminopeptidase N,APN)多肽等对不同的Cry毒素也具有协同增效作用[10]。原核表达的钙粘蛋白CR12-MPED片段通过增加Cry1A在昆虫中肠BBMV上的结合位点,吸引Cry1A与中肠上皮细胞膜结合,进而显著增强了Cry1A 对烟草天蛾(Manducasexta)、烟芽夜蛾(Helicoverpavirescens)和美洲棉铃虫(HelicoverpazeaBoddie)幼虫的毒性[32]。研究表明,烟草天蛾中肠钙粘蛋白的CR7和CR11区域片段同样可增强Cry1Ac和Cry1Ab对烟草天蛾幼虫的毒性,并进一步证实钙粘蛋白片段能促使毒素的寡聚化,从而对毒素起到增效作用[24]。玉米根叶甲钙粘蛋白的CR 8~10区域片段与Cry3Aa、Cry3Bb和Cry8Ca毒素结合,有利于Cry毒素寡聚化,进而插入膜内以实现其增效作用[33]。草地夜蛾(Spodopterafrugiperda)和烟草天蛾的钙粘蛋白(SfCad与MsCad)通过保护Cry1Fa免受昆虫中肠蛋白酶的降解,提高了Cry1Fa在其中肠上的聚集,增加了Cry1Fa的杀虫活性[25]。原核表达的冈比亚按蚊 (Anophelesgambiae)28 kDa AgAPN2ta片段和30 kDa AgAPN2tb片段与Bt Cry11Ba毒素的结合显示出不同的协同作用,即AgAPN2tb片段显著增强了Cry11Ba毒素的毒性,而AgAPN2ta片段却较大程度的抑制了Cry11Ba毒素的毒性[10]。此外,一些重组表达的Bt毒素蛋白和钙粘蛋白之间也可实现协同增效作用,如Cry11Ba与Cry4Aa、Cry46Ab与Cry4Aa对尖音库蚊(Culexpipiens)幼虫[34,35],重组钙粘蛋白(SlABCC3和Hacadherin)与活化的Cry1Ac对斜纹夜蛾(Spodopteralitura)幼虫[36],Cyt1Aa与Cry4Aa对蚋属蝇(Simuliumspp.)幼虫的毒性等[37]。
2.4 降低目标昆虫中肠完整性,加快毒素蛋白作用的发挥
围食膜(peritrophic membrane,PM)主要由几丁质和蛋白质组成,可保护中肠上皮细胞,阻止病原物等侵染,是中肠细胞和肠腔之间的防御性屏障[38]。完整围食膜中几丁质、酸性粘多糖、粘蛋白(intestinal mucin,IM),以及其他多种蛋白质的降解消化,将使围食膜的致密性、坚韧性降低,渗透性增强,从而使Bt和病原微生物的通过率提高,进而实现对Bt的增效作用[39]。病原性病毒的增效蛋白或蛋白因子等通过对围食膜组成的降解,使其结构改变,通透性增加。如经口感染昆虫的核型多角体病毒(nuclear polyhedrosis virus, NPV)后,Bt更加顺利地通过围食膜到达中肠细胞,进而使其对昆虫的感染力和自身利用效率得以提高,从而实现对Bt毒力的增效活性[40]。其次,使完整性围食膜难以生成也是实现对Bt增效的途径之一。通过与中肠新合成的几丁质结合,或者阻止围食膜蛋白与几丁质的正常结合、昆虫中肠几丁质微纤丝的形成,进而导致完整围食膜难以形成,从而实现毒素蛋白和病原微生物的高效通过率[39,41]。
2.5 改善环境因素,降低毒素蛋白杀虫活性的损失
自然辐射直接或通过水介质中产生的自由基和活性氧群的间接作用对机体内DNA或蛋白质等细胞内生物大分子造成损伤,使其生物活性降低或丧失。同样,ICPs、α-外毒素、β-外毒素、γ-外毒素、VIPs和芽孢等也受高温、强光、紫外辐射和电离辐射等环境因素的影响,进而影响其杀虫活性。即在施用的自然环境中受多种环境因素的影响(包括温度、光照、紫外辐射和电离辐射等),原毒素的组氨酸和酪氨酸等组成氨基酸受到损伤,多级结构均被破坏,进而变性,难以溶解和激活成具有杀虫活性的毒素蛋白,从而与靶昆虫中肠受体的结合受到影响,进而降低杀虫活性[2,25,41,42]。黑色素、二苯乙烯类荧光增白剂等光稳定剂或其他作为Bt制剂增效剂的增效机制是,通过反射可见光,吸收高能量紫外辐射;或者通过提供质子,捕获自由基,形成稳定的共振结构,阻断氧化;或者通过与氧作用消耗密闭系统中残留的氧;或者通过螯合金属离子,减少金属离子对氧化的催化作用;或者通过淬灭单线态氧,阻止光氧化[41,43]。
2.6 其他
Bt蛋白的杀虫活性得以实现,需经过摄食、溶解、酶解激活、结合受体、插入上皮细胞,以及形成孔洞或离子通道等作用过程,修饰、改变其中任何环节都会对Bt蛋白的杀虫活性产生影响。然而,除上述作用途径或方式外,增效剂通过延缓昆虫对毒素蛋白产生抗性或改变昆虫体内生化途径等也可实现对Bt制剂的增效作用。如纳米银颗粒通过延缓埃及伊蚊对Bt以色列菌株产生抗性,以达到协同增效作用[43]。马来酸与柠檬酸等通过参与三羧酸循环,增强了昆虫体内相关酶的活性,进而使得昆虫对杀虫毒素变得更为敏感,从而实现其增效作用[44]。而植物提取物则通过改变Bt制剂的物理性状、干扰昆虫的生理生化指标,或者通过目标昆虫自身存在的感觉和/或免疫系统,对自身行为进行调节,从而实现其增效作用[45~49]。
3 展望
作为研究较为深入的生物杀虫剂,Bt毒素蛋白的种类、氨基酸残基序列、杀虫谱和三维结构等均具有多样性,其不同于常规的化学杀虫剂,与昆虫靶标之间的相互作用已进化到极其复杂的水平。毒素蛋白杀虫活性的发挥需要多种因素的共同作用,同时,昆虫对毒素蛋白的反应也具有多样性,因此,Bt杀虫机制与增效作用机理的不确定性需要进一步的揭示,且需要多方位的进行。分子生物学是目前研究中比较集中采用的手段或途径,且主要集中于研究不同毒素蛋白之间的协同作用。然而,Bt杀虫活性的实现是一系列的系统过程(包括孔洞模式和信号转导模式),也需要考虑目标害虫的生理生化变化、组织等的病理学改变与摄食毒素蛋白(增效剂)后的改变等内在因素。其次,除内在因素外,昆虫的化学感觉器作用、寄主植物与增效剂间的化感作用等外在变化也需要进一步的深入研究。最后,研究还发现寡聚化和插入是Cyt1Aa实现杀虫活性的必要条件,但不是充分条件[11],因此,Bt毒素蛋白的杀虫活性机制是否存在限速步骤,若存在,又能否通过限制性步骤进一步实现增效剂的增效活性,这为深度了解晶体毒素的杀虫和增效机制提出了新的研究思路。
[1] Schnepf E, Crickmore N, Van Rie J,etal..Bacillusthuringiensisand its pesticidal crystal proteins[J]. Microbiol. Mol. Biol. R., 1998, 62(3): 775-806.
[2] 申继忠,钱传范. 苏云金杆菌杀虫剂增效途径研究[J]. 生物防治通报,1994, 10(3): 135-140.
[3] 金大勇, 李翔国, 张吉子. 苏云金杆菌制剂的增效因子研究进展[J]. 延边大学农学学报, 2012, 34(3):267-272.
[4] Aronson A I, Shai Y. WhyBacillusthuringiensisinsecticidal toxins are so effective: Unique features of their mode of action[J]. FEMS Microbiol. Lett., 2001, 195: 1-8.
[5] Gill S S, Cowles E A, Pietrantonio P V. The mode of action ofBacillusthuringiensisendotoxin[J]. Ann. Rev. Entomol., 1992, 37: 615-636.
[6] Pérez C, Munoz-Garay C, Portugal L C,etal..Bacillusthuringiensissp.israelensisCyt1Aa enhances activity of Cry11Aa toxin by facilitating the formation of a pre-pore oligomeric structure[J]. Cell Microbiol., 2007, 9(12): 2931-2937.
[7] Zhang X B, Candas M, Griko N B,etal.. Cytotoxicity ofBacillusthuringiensisCry1Ab toxin depends on specific binding of the toxin to the cadherin receptor BT-R1expressed in insect cells[J]. Cell Death Differ., 2005, 12(11): 1407-1416.
[8] Sangadala S, Walters F S, English L H,etal.. A mixture ofManducasextaaminopeptidase and phosphatase enhancesBacillusthuringiensisinsecticidal Cry1A(c) toxin binding and86Rb+-K+effluxinvitro[J]. J. Biol. Chem., 1994, 269(13): 10088-10092.
[9] Zhang X B, Candas M, Griko N B,etal.. A mechanism of cell death involving an adenylyl cyclase/PKA signaling pathway is induced by the Cry1Ab toxin ofBacillusthuringiensis[J]. Proc. Natl. Acad. Sci. USA, 2006, 103(26): 9897-9902.
[10] 陈文波,刘晨曦,吴孔明. Cry毒素昆虫中肠受体蛋白多肽的增效作用研究进展[J]. 中国生物防治学报, 2013, 29(1): 117-123.
[11] Tanaka S, Yoshizawa Y, Sato R. Response of midgut epithelial cells to Cry1Aa is toxin-dependent and depends on the interplay between toxic action and the host apoptotic response[J]. FEBS J., 2012, 279(6): 1071-1079.
[12] Barbieri R, Cavallini G, Malavolta C,etal.. Comparison of the effectiveness of different commercial products based onBacillusthuringiensisBerliner againstLobesiabotrana(Den and Schiff.) [J]. Inform. Fitopatdogico, 1988, 38: 7-8,59-62.
[13] Zhang J H, Wang C Z, Qin J D. Effect of feeding stimulant on the feeding behavior and mortality ofHelicoverpaarmigeraon diets withBacillusthuringiensis[J]. Entomol. Sin., 2000, 7(2): 155-160.
[14] 邱占奎,袁会珠,李永平,等.添加剂有机硅表面活性剂对低容量喷雾防治小麦蚜虫的影响[J]. 植物保护,2006, 32(2): 34-36.
[15] El-Moursy A, Aboul-Ela R, Salama H S,etal.. Chemical additives that effect the potency of endotoxin ofBacillusthuringiensisagainstPlodiainterpunctella[J]. Int. J. Trop. Insect Sci., 1992, 13(6): 775-779.
[16] 严东辉, 陈昌洁.提高昆虫病毒杀虫效果的荧光增白剂的研究[J]. 林业科学,2003, 39(1): 153-159.
[17] Morris O N, Converse V, Kanagaratnam P. Chemical additive effects on the efficacy ofBacillusthuringiensisBerliner subsp.KurstakiagainstMamestraconfigurata(Lepidoptera: Noctuidae)[J]. J. Econ. Entomol., 1995, 88(4): 815-824.
[18] Salama H S, Abd El-Baset M S, Ragaei M. Mode of action of chemical additives in enhancing the potency ofBacillusthuringiensisagainst lepidopterous insects[J]. J. Appl. Entomol., 1992, 114(2): 167-173.
[19] Juntadech T, Kanintronkul Y, Kanchanawarin C,etal.. Importance of polarity of the α4-α5 loop residue——Asn166in the pore-forming domain of theBacillusthuringiensisCry4Ba toxin: Implications for ion permeation and pore opening[J]. Biochim. Biophys. Acta, 2014, 1838: 319-327.
[20] Kaikaew A, Promptmas C, Angsuthanasombat C. Importance of Thr328and Thr369for functional maintenance of two receptor-binding β-hairpins of theBacillusthuringiensisCry4Ba toxin: Implications for synergistic interactions with Cyt2Aa2[J]. Biochem. Bioph. Res. Co., 2016, 469: 698-703.
[21] Cantón P E, Reyes E Z, De Escudero I R,etal.. Binding ofBacillusthuringiensissubsp.israelensisCry4Ba to Cyt1Aa has an important role in synergism[J]. Peptides, 2011, 32: 595-600.
[22] Lailak C, Khaokhiew T, Promptmas C,etal..BacillusthuringiensisCry4Ba toxin employs two receptor-binding loops for synergistic interactions with Cyt2Aa2[J]. Biochem. Bioph. Res. Co., 2013, 435: 216-221.
[23] Elleuch J, Jaoua S, Darriet F,etal.. Cry4Ba and Cyt1Aa proteins fromBacillusthuringiensisisraelensis: Interactions and toxicity mechanism againstAedesaegypti[J]. Toxicon, 2015, 104: 83-90.
[24] Pacheco S, Gómez I, Gill S S,etal.. Enhancement of insecticidal activity ofBacillusthuringiensisCry1A toxins by fragments of a toxin-binding cadherin correlates with oligomer formation[J]. Peptides, 2009, 30: 583-588.
[25] Rahman K, Abdullah M A F, Ambati S,etal.. Differential protection of Cry1Fa toxin againstSpodopterafrugiperdalarval gut proteases by cadherin orthologs correlates with increased synergism[J]. Appl. Environ. Microbiol., 2011, 78(2): 354-362.
[26] Xue J L, Cai Q X, Zheng D S,etal.. The synergistic activity between Cry1Aa and Crylc fromBacillusthuringiensisagainstSpodopteraexiguaandHelicoverpaarmigera[J]. Lett. Appl. Microbiol., 2005, 40(6): 460-465.
[27] 杨庆仙.苏云金杆菌增效细菌研究进展[J]. 北方园艺,2008, 1: 55-58.
[28] Wraight S P, Ramos M E. Characterization of the synergistic interaction betweenBeauveriabassianastrain GHA andBacillusthuringiensismorrisonistrain tenebrionis applied against Colorado potato beetle larvae[J]. J. Invertebr. Pathol., 2017, 144: 47-57.
[29] Baranek J, Konecka E, Kaznowski A. Interaction between toxin crystals and vegetative insecticidal proteins ofBacillusthuringiensisin lepidopteran larvae[J]. BioControl, 2017, 62: 649-658.
[30] Cantón P E, López-Díaz J A, Gill S S,etal.. Membrane binding and oligomer membrane insertion are necessary but insufficient forBacillusthuringiensisCyt1Aa toxicity[J]. Peptides, 2014, 53: 286-291.
[31] Tabashnik B E. Evaluation of synergism amongBacillusthuringiensistoxins[J]. Appl. Environ. Microbiol., 1992, 58(10): 3343-3346.
[32] Chen J, Hua G, Jurat-Fuentes J L,etal.. Synergism ofBacillusthuringiensistoxins by a fragment of a toxin-binding cadherin[J]. Proc. Natl. Acad. Sci. USA, 2007, 104: 13901-13906.
[33] Park Y J, Hua G, Taylor M D,etal.. A coleopteran cadherin fragment synergizes toxicity ofBacillusthuringiensistoxins Cry3Aa, Cry3Bb, and Cry8Ca against lesser mealworm,Alphitobiusdiaperinus(Coleoptera: Tenebrionidae)[J]. J. Invertebr. Pathol., 2014, 123: 1-5.
[34] Hayakawa T, Yoneda N, Okada K,etal..BacillusthuringiensisCry11Ba works synergistically with Cry4Aa but not with Cry11Aa for toxicity against mosquitoCulexpipiens(Diptera: Culicidae)larvae[J]. Appl. Entomol. Zool., 2017, 52: 61-68.
[35] Hayakawa T, Sakakibara A, Ueda S,etal.. Cry46Ab fromBacillusthuringiensisTK-E6 is a new mosquitocidal toxin with aerolysin-type architecture[J]. Insect Biochem. Molec., 2017, 87: 100-106.
[36] Chen Z W, He F, Xiao Y T,etal.. Endogenous expression of a Bt toxin receptor in the Cry1Ac-susceptible insect cell line and its synergistic effect with cadherin on cytotoxicity of activated Cry1Ac[J]. Insect Biochem. Molec., 2015, 59: 1-17.
[37] Monnerat R, Pereira E, Teles B,etal.. Synergistic activity ofBacillusthuringiensistoxins againstSimuliumspp. larvae[J]. J. Invertebr. Pathol., 2014, 121: 70-73.
[38] Hegedus D, Erlandson M, Gillott C,etal.. New insights into peritrophic matrix synthesis, atchitecture, and function[J]. Annu. Rev. Entomol., 2009, 54: 285-302.
[39] Wang P, Granados R R. Observations on the presence of the peritrophic membrane in larvalTrichoplusianiand its role in limiting baculovirus infection[J]. J. Invertebr. Pathol., 1998, 72: 57-62.
[40] 徐 健,刘 琴,祝树德.昆虫颗粒体病毒增效蛋白研究进展[J]. 华东昆虫学报, 2005, 14(4): 343-347.
[41] 徐 莉,杨 红,彭建新,等.荧光增白剂对苏云金芽孢杆菌毒力的增效作用及其紫外防护功效[J]. 中国生物防治,2001, 17(2): 63-66.
[42] 王文军,钱传范,申继忠,等.活性氧对苏云金芽孢杆菌伴孢晶体的损伤作用[J]. 微生物学报,1999, 39(5): 469-474.
[43] Saxena D, Ben-Dov E, Manasherob R,etal.. A UV tolerant mutant ofBacillusthuringiensissubsp.kurstakiproducing melanin[J]. Curr. Microbiol., 2002, 44: 25-30.
[44] Narayanan K. Insect defense: Its impact on microbial control of insect pests[J]. Curr. Sci., 2004, 86(6): 800-814.
[45] 魏 辉,王前梁,侯有明,等.植物提取物对苏云金芽孢杆菌杀虫活性的影响[J]. 农药学学报,2003, 5(4): 75-79.
[46] 魏 辉,杨 广,王前梁,等.小菜蛾幼虫头部化学感觉器的电镜扫描[J]. 福建农林大学学报(自然科学版),2003, 32(4): 434-437.
[47] Van Loon J J, Van L. Chemosensory basis of feeding and oviposition behaviour in herbivorous insects: A glance at the periphery[J]. Entomol. Exp. Appl., 1996, 80: 7-13.
[48] Yaroslavtseva O N., Dubovskiy I M, Khodyrev V P,etal.. Immunological mechanisms of synergy between fungusMetarhiziumrobertsiiand bacteriaBacillusthuringiensisssp.morrisonion Colorado potato beetle larvae[J]. J. Insect Physiol., 2017, 96: 14-20.
[49] Zhou W B, Orr M W, Lee V T,etal.. Synergistic effects of ultrafast heating and gaseous chlorine on the neutralization of bacterial spores[J]. Chem. Eng. Sci., 2016, 144: 39-47.
