藤黄酸对耐药胃癌细胞增殖、凋亡、迁移的影响及机制
2017-05-03丁忠阳许飞唐建东李淦蒋盘强吴昊天唐张峰
丁忠阳,许飞,唐建东,李淦,蒋盘强,吴昊天,唐张峰
(南京中医药大学附属无锡市中医医院,江苏无锡214071)
藤黄酸对耐药胃癌细胞增殖、凋亡、迁移的影响及机制
丁忠阳,许飞,唐建东,李淦,蒋盘强,吴昊天,唐张峰
(南京中医药大学附属无锡市中医医院,江苏无锡214071)
目的 探讨藤黄酸对耐药胃癌细胞增殖、凋亡、迁移的影响及机制。方法 取处于对数期的耐吉西他滨的胃癌细胞(R-MKN28),分为未处理组(Blank组)、藤黄酸组和藤黄酸+IL-6组,Blank组不予任何干预,藤黄酸组予藤黄酸2.0 μg/mL,藤黄酸+IL-6组在藤黄酸处理的基础上加用IL-6 10 ng/mL 处理。用MTT、流式细胞术、Transwell和生物信息学等方法观察藤黄酸对R-MKN28细胞增殖、凋亡和迁移的影响。并通过qRT-PCR法检测IL-6、JAK1、BCL2和AKT1基因表达,采用ELISA法检测IL-6蛋白表达。结果 藤黄酸干预至72 h时,Blank组492 nm处吸光度值由0 h的0.472±0.012升高到0.988±0.031,而藤黄酸组只由0 h的0.409±0.008上升至0.762±0.022,两组比较,P<0.01。细胞凋亡检测结果显示,Blank组凋亡率为(14.25±1.352)%;藤黄酸组凋亡率为(23.48±1.574)%,两组比较,P<0.05。Transwell 结果显示,Blank组穿膜细胞数为(61.67±7.265)个/视野,而藤黄酸组穿膜细胞数为(29.68±5.487)个/视野,两组比较,P<0.05。藤黄酸组R-MKN28细胞的IL-6、JAK1、BCL2和AKT1基因表达水平表达下降,IL-6蛋白表达减少(P均<0.05)。MTT实验结果显示,藤黄酸+IL-6组细胞增殖能力相比藤黄酸组在48 h后明显上升(P<0.05)。Transwell 实验结果显示,藤黄酸+IL-6组穿膜细胞数为(60.00±4.619)个/视野,而藤黄酸组仅为(27.00±5.859)个/视野,两组比较,P<0.05。生物信息学分析表明耐吉西他滨的胃癌细胞中IL-6水平表达升高。结论 藤黄酸可能通过调节IL-6水平抑制耐吉西他滨的 R-MKN28 细胞增殖和迁移,促进凋亡。
胃癌;耐药癌细胞;藤黄酸;白细胞介素6
胃癌是世界范围内最常见的恶性肿瘤之一[1],发病率逐年上升,病死率较高。外科手术是目前最有效的治疗方法,但我国胃癌的早期诊断率低,确诊时大部分患者已失去手术机会;另外,50%~70%进展期胃癌患者术后可出现复发[2]。化疗是一种全身性治疗手段,对潜在转移灶和临床已发生转移的癌灶均有作用。因此,化疗已成为患者术后辅助性治疗和晚期不适于手术患者姑息治疗的主要方式。但随着化疗药物的持续治疗,患者机体可产生不同程度的耐药导致化疗效果下降甚至无效[3]。在我国,中医药治疗癌症的方式被广泛采用,且取得较好效果[4]。2014年9月,我们采用藤黄酸中药单体处理胃癌耐药细胞,观察藤黄酸对胃癌耐药细胞耐吉西他滨的作用,并对其机制进行研究。
1 材料与方法
1.1 材料 藤黄酸粉剂(纯度99%)和MTT试剂购自美国Sigma 公司,溶解分装后保存于-20 ℃。细胞培养相关试剂购于美国Gibco公司;细胞培养材料购于美国Corning公司。人胃癌耐吉西他滨细胞系R-MKN28由Saiki Y 教授馈赠,R-MKN28 细胞培养于含10% FBS和双抗(100 U/mL青霉素、50 mg/mL链霉素)的高糖DMEM培养基中,置于37 ℃含5% CO2孵箱中。TRIzol购于Invitrogen公司。PCR相关试剂购于大连宝生生物公司。IL-6的ELISA试剂盒购于北京百奥莱博科技有限公司。
1.2 细胞增殖能力检测 取对数期R-MKN28细胞传代接种于96孔板(1×103个/孔)。待细胞贴壁后,分为未处理组(Blank组)、藤黄酸组,Blank组不予任何干预,藤黄酸组加入100 μL溶解的藤黄酸溶液,使藤黄酸终浓度为2.0 μg/mL,此时记为0 h。继续培养细胞,分别于0、24、48 h和72 h加入20 μL/孔MTT试剂,放入孵箱再培养4 h,取出96孔板,用注射器小心吸走细胞培养基,加入150 μL二甲基亚砜(DMSO),避光振摇10 min,上机于492 nm 波长处检测每孔吸光度值(OD值)。
1.3 细胞凋亡率检测 取对数期R-MKN28细胞接种于6孔板,细胞数为8×105个/孔。12 h细胞贴壁后,分为未处理组(Blank组)、藤黄酸组,Blank组不予任何干预,藤黄酸组加入藤黄酸溶液,使其终浓度为2.0 μg/mL。培养48 h后将细胞用胰酶消化后收集至15 mL离心管,保证每个样本细胞数≥106。用预冷PBS洗涤细胞3次。加入 Anexin V 和 PI 进行孵育(注意避光)。送实验中心流式平台检测细胞凋亡率。
1.4 细胞迁移能力检测 R-MKN28细胞接种于6孔板,细胞贴壁后分为未处理组(Blank组)、藤黄酸组,Blank组不予任何干预,藤黄酸组加入藤黄酸2.0 μg/mL进行干预。24 h后将细胞胰酶消化后收集于离心管,1 000 r/min,离心3 min。PBS洗涤1次,加入无血清培养基重悬。细胞计数后,将400 μL共5×105个细胞加入悬挂于24孔板上的Transwell小室上室。下室为20% FBS培养基600 μL。24 h后取出Transwell小室,PBS洗涤2次,加入结晶紫进行染色。镜下观察每个小室细胞数目,并计算每个视野下细胞数目。
1.5 R-MKN28细胞中IL-6、JAK1、BCL2和AKT1基因表达检测 采用实时荧光定量PCR法。R-MKN28 细胞分为未处理组(Blank组)、藤黄酸组,Blank组不予任何干预,藤黄酸组加入藤黄酸2.0 μg/mL进行干预。36 h后使用TRIzol提取细胞RNA。取2 μg逆转录为cDNA。按qRT-PCR试剂盒操作说明,依次加入模板、引物和预混SYBGREEN 等试剂进行 qRT-PCR 反应检测基因表达,依照2-ΔΔCT计算。引物序列IL-6上游引物5′-CTCAATATTAGAGTCTCAACCCCCA-3′,下游引物5′-GAGAAGGCAACTGGACCGAA-3′;JAK1上游引物5′-GCATCGAGCGCACAAAGTTAT-3′,下游引物5′-ACCAGTAGGGTTGAGGGACA-3′;BCL2上游引物5′-GGGAGGATTGTGGCCTTCTT-3′,下游引物5′-ACTTGTGGCCCAGATAGGCA-3′;AKT1上游引物5′-GAAGGACGGGAGCAGGCG-3′,下游引物5′-TGTACTCCCCTCGTTTGTGC-3′。
1.6 IL-6蛋白表达检测 采用ELISA法。严格按照试剂盒说明书进行操作,对数生长期的 R-MKN28细胞接种于 6 孔板,每孔细胞数为5×106个。待细胞贴壁后分为未处理组(Blank组)、藤黄酸组,Blank组不予任何干预,藤黄酸组加入终浓度为2.0 μg/mL的藤黄酸,继续培养24 h,之后加入裂解液进行蛋白抽提,抽提完成后加入已包被的板子,加入200 μL抗IL-6的酶标抗体,室温放置2 h。洗涤液清洗4 次,每孔加200 μL 底物进行显色。室温反应20 min,每孔加入50 μL反应终止液,采用酶标仪在450 nm 处检测。
1.7 生物信息学分析 芯片数据来源为GEO数据平台上 GEO accession 为 GSE58118 数据 (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE58118)。该基因表达谱芯片数据包含4个样本,其中2个R-MKN28细胞样本,2 个耐吉西他滨的R-MKN28细胞样本。芯片数据下载后对数据整理,使用斯坦福大学开发的SAM软件对芯片数据分析,采用heatmap illustrator软件导出热图。
1.8 藤黄酸处理R-MKN28后加入IL-6重组蛋白对细胞增殖和迁移能力的影响 取对数生长期的 R-MKN28细胞接种于 6 孔板,每孔细胞数为5×106个。待细胞贴壁后分为未处理组(Blank组)、藤黄酸组、藤黄酸+IL-6组,Blank组不予任何干预,藤黄酸组加入终浓度为2.0 μg/mL的藤黄酸,藤黄酸+IL-6组予藤黄酸2.0 μg/mL及重组IL-6蛋白10 ng/mL,细胞增殖能力检测同1.2,细胞迁移能力检测同1.4。
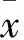
2 结果
2.1 藤黄酸对R-MKN28细胞增殖、凋亡及迁移的影响 加入藤黄酸后R-MKN28细胞增殖于24 h后明显降低(P<0.05),且至72 h此现象更明显(P<0.01);R-MKN28细胞凋亡增加(P<0.05);藤黄酸组R-MKN28 细胞穿膜数数明显低于对照组(P<0.05),见表1。
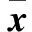
表1 两组细胞增殖、凋亡和迁移能力比较(±s)
2.2 藤黄酸处理后R-MKN28 细胞相关基因表达变化 qRT-PCR 结果显示,加入藤黄酸后 R-MKN28细胞IL-6基因表达水平相比Blank组的4.333下降至1.167、JAK1相比Blank组的 2.786下降至1.133、BCL2相比Blank组的3.300下降至1.000和AKT1相比Blank组的3.300下降至0.933;藤黄酸组IL-6 蛋白表达(111.333 ng/mL)相比Blank组的(169.000 ng/mL)亦下降(P均<0.05)。
2.3 耐吉西他滨胃癌的生物信息学分析 下载 GEO 数据库GEO accession 为 GSE58118 的基因芯片进行分析表明,耐吉西他滨胃癌细胞相比不耐药细胞株IL-6基因水平表达明显增高。
2.4 藤黄酸处理R-MKN28 后恢复IL-6水平对R-MKN28细胞增殖和迁移能力的影响 加入重组IL-6后R-MKN28细胞增殖和迁移能力均有不同程度恢复(P均<0.05),见表2。
表2 各组细胞在恢复IL-6水平后增殖和迁移能力比较(±s)
3 讨论
晚期胃癌是目前外科临床研究的重点、难点。尤其是经过多重化疗后,部分患者出现对常规化疗方案的耐药,治疗棘手[5]。吉西他滨是一种人工合成的嘧啶类核苷药物,其主要作用机制是干扰细胞NDA周期,为周期特异性药物。吉西他滨目前已大范围用于食管癌、乳腺癌和原发性肝癌等多种肿瘤的治疗[6]。关于吉西他滨治疗胃癌的文献较多,其中多数研究认为吉西他滨对胃癌无明显作用[7]。
藤黄酸为藤黄科植物分泌的干燥树脂藤黄中主要成分。中医古籍记载藤黄具有消炎、抑菌、止血、抗病毒等作用,常被用于脓肿、跌打损伤和各种体藓的治疗中。藤黄酸是橙黄色无定形粉末,具有光学性,相对分子质量为628.76 kD。药理学研究表明藤黄酸具有抗肿瘤作用,可抑制多种癌症的增殖、迁移、侵袭。藤黄酸可通过线粒体氧化应激和诱导细胞色素C的释放来促进鼻咽癌细胞的凋亡[8],通过将肺癌细胞周期阻滞于G0/G1期达到抑制肿瘤细胞增殖的作用[9]。本研究结果显示,耐吉西他滨胃癌细胞R-MKN28加入藤黄酸后,细胞增殖明显下降,且呈时间依赖性;细胞凋亡增加;细胞迁移能力降低。可见藤黄酸可影响耐吉西他滨的胃癌细胞生物学行为,抑制耐吉西他滨的胃癌细胞生长并促进凋亡。本研究通过qRT-PCR检测发现藤黄酸可降低R-MKN28细胞IL-6基因和蛋白及其下游基因水平。IL-6是一种可由多种细胞分泌的多功能因子。在多种肿瘤的宿主防御和癌细胞生长中起中心作用。在胃癌中,IL-6可通过激活c-Src/RhoA/ROCK信号通路介导胃癌的侵袭[10]。IL-6/STAT3/Jagged-1/Notch 信号通路是曲妥单抗治疗胃癌发挥作用的重要信号通路,亦是胃癌耐曲妥单抗的主要靶点[11]。生物信息学分析结果表明在耐吉西他滨的胃癌细胞中IL-6表达水平明显升高,而加入藤黄酸后IL-6表达水平下降,且在使用重组IL-6后R-MKN28生物学行为又朝恶性程度方向发展。
综上所述,虽然多数文献报道吉西他滨不能用于胃癌治疗,但此研究中我们加入了中药单体藤黄酸后,胃癌细胞增殖、迁移能力下降,凋亡增多。此可能成为吉西他滨治疗胃癌的一个突破口,但具体机制尚需深入研究。
[1] Pecqueux M, Fritzmann J, Adamu M, et al. Free intraperitoneal tumor cells and outcome in gastric cancer patients: A systematic review and meta-analysis [J]. Oncotarget, 2015,6(34):35564-35578.
[2] Iacovelli R, Pietrantonio F, Maggi C, et al. Combination or single-agent chemotherapy as adjuvant treatment of gastric cancer: A systematic review and meta-analysis of published trials [J]. Crit Rev Oncol Hematol, 2016,(98):24-28.
[3] Wu C, Zhuang Y, Jiang S, et al. Interaction between wnt/β-catenin pathway and micrornas regulates epithelial-mesenchymal transition in gastric cancer (review) [J]. Int J Oncol, 2016,48(6):2236-2246.
[4] Qu Z, Zou X, Zhang X, et al. Chelidonine induces mitotic slippage and apoptotic-like death in sgc-7901 human gastric carcinoma cells [J]. Mol Med Rep, 2016,13(2):1336-1344.
[5] Zhang G, Wang Z, Qian F, et al. Silencing of the abcc4 gene by rna interference reverses multidrug resistance in human gastric cancer [J]. Oncol Rep, 2015,33(3):1147-1154.
[6] Gaddy DF, Becker C, Li H, et al. A systematic literature review to identify and compare clinical trials evaluating novel therapeutic agents in post-gemcitabine advanced pancreatic cancer[J]. Value Health, 2015,18(7):A434.
[7] Okusaka T, Ishii H, Funakoshi A, et al. Phase ii study of single-agent gemcitabine in patients with advanced biliary tract cancer [J]. Cancer Chemother Pharmacol, 2006,57(5):647-653.
[8] Yan F, Wang M, Chen H, et al. Gambogenic acid mediated apoptosis through the mitochondrial oxidative stress and inactivation of akt signaling pathway in human nasopharyngeal carcinoma cne-1 cells[J]. Eur J Pharmacol, 2011,652(1):23-32.
[9] Li Q, Cheng H, Zhu G, et al. Gambogenic acid inhibits proliferation of a549 cells through apoptosis-inducing and cell cycle arresting [J]. Biol Pharm Bull, 2010,33(3):415-420.
[10] Lin MT, Lin BR, Chang CC, et al. IL-6 induces ags gastric cancer cell invasion via activation of the c‐src/rhoa/rock signaling pathway [J]. Int J Cancer, 2007,120(12):2600-2608.
[11] Yang Z, Guo L, Liu D, et al. Acquisition of resistance to trastuzumab in gastric cancer cells is associated with activation of il-6/stat3/jagged-1/notch positive feedback loop[J]. Oncotarget, 2015,6(7):5072-5087.
2015年度无锡市科技发展资金项目(CSE31N1513)。
10.3969/j.issn.1002-266X.2017.09.012
R735.2
A
1002-266X(2017)09-0039-03
2016-07-01)
