水生动物肌醇营养研究进展
2015-12-21杨奇慧谭北平董晓慧迟淑艳刘泓宇
杨 杰 杨奇慧 谭北平 董晓慧 迟淑艳 刘泓宇 章 双
(广东海洋大学水产学院,水产动物营养与饲料实验室,湛江 524088)
肌醇是环己六醇族的六羰基环己烷,即环己六醇,俗名为心肌糖,属水溶性维生素类产品,性质稳定。肌醇分子式为C6H12O6,相对分子质量是180.16,它共有9个异构体,其中仅1种具生物活性形式,即内消旋(1,2,3,5-顺式)环己醇。具有生物活性的肌醇是白色结晶或结晶状粉末,溶于水但不溶于醇和醚[1-3]。
肌醇与大多数水溶性维生素不同,在体内不是以辅酶的形式参与代谢,而是作为体内活性组织的组成部分发挥功能,其主要有以下几种生理功能:1)肌醇是动物体内磷脂结构组织的主要组成部分,以磷脂酰肌醇的形式参与膜结构,维持细胞正常结构和功能。2)肌醇可以通过异生途径生成糖类供能。3)肌醇作为环已烷的衍生物,具有很明显的亲脂性,可防止体内胆固醇的积累,预防脂肪肝,与胆碱一起参与脂肪代谢的平衡调控[2]。
1 水生动物肌醇缺乏症
肌醇主要以六磷酸盐的形式存在于谷物饲料中,陆生动物肝脏、肾脏、睾丸和其他组织的肌醇合成量足以满足其需要量,在饲料中不需要添加。但以豆科类及其加工副产品为主要蛋白质原料的水生动物饲料,由于谷物饲料的用量很少和自身合成能力的限制,故必需添加肌醇[4]。
目前已经在建鲤(Cyprinus carpio var.Jian)[3-4]、斑点叉尾 (Ictalurus punctatus)[5]、奥尼罗非鱼(Oreochromis niloticus×Oreochromis aureus)[6-7]、尼罗罗非鱼(Oreochromis niloticus)[8]、草鱼(Ctenopharyngodon idella)[9]、大西洋鲑(Salmo salar L.)[10]、军 曹 鱼 (Rachycentron canadum)[11]、大 黄 鱼 (Poseudosciaena crocea)[12]、鲈(Lateolabrax japonicas)[12]、牙鲆(Paralichthys olivaceus)[13]、卵形鲳鲹(Trachinotus ovatus)[14]、斑节对虾(Penaeus monodon)[15]、中华绒螯蟹(Eriocheir sinensis)[16]、皱纹盘鲍(Haliotis discus hannai Ino)[17]、黑鲷(Sparus marcocephalus)[18]等水生动物上开展了肌醇缺乏的研究。研究主要集中在饲料中肌醇缺乏对水生动物生长性能的影响上。研究表明:水生动物在饲料中肌醇缺乏时,表现出明显的肌醇缺乏症;水生动物的种类和生理阶段不同,肌醇缺乏的症状也存在差异。水生动物肌醇缺乏的总体表现是:生长不良、皮肤出血、鳍条糜烂、体色发黑、脂肪肝等症状(表1)。水生动物饲料中肌醇缺乏所表现出来的典型症状与肌醇在其体内的生理功能关系密切,本文将通过以下3个方面对肌醇的营养生理作用进行简述。

表1 水生动物肌醇缺乏的典型症状Table 1 Typical deficiency syndromes of myo-inositol in aquatic animals
1.1 肌醇与生长
已有研究显示,肌醇主要通过影响摄食量和饲料效率改善水生动物的生长性能。肌醇缺乏时导致黑鲷幼鱼胃胀,空胃时间长[18]。这可能是因为肌醇加速了水生动物的胃肠排空,缩短了饥饿时间,从而促进了摄食[4]。肌醇加速胃肠排空的可能途径:1)肌醇提高了肠道消化吸收能力,使消化吸收加速,从而加速了胃肠排空,促使动物摄食。姜维丹[3]对建鲤的研究显示,肌醇显著提高了幼建鲤肝胰脏中的胰蛋白酶、糜蛋白酶、脂肪酶和淀粉酶、肠道中的胰蛋白酶、糜蛋白酶、脂肪酶和淀粉酶活力;张美彦等[19]研究发现,饲料中添加肌醇能够促进幽门盲囊和肠道中蛋白酶和脂肪酶的分泌,从而有助于哲罗鲑(Hucho taimen)对蛋白质和脂肪的分解,提高其对饲料的消化吸收能力。2)肌醇可能间接调控了胃肠道的收缩反应。Peppiatt等[20]研究发现,肌醇可以通过与第二信使系统的交互作用,最终调控细胞质中的钙离子浓度来影响神经活动,上调钙离子的释放,从而促进了乙酰胆碱的释放,通过胆碱能神经途径促进胃肠道的收缩反应。当肌醇缺乏时,这种收缩反应失调,导致胃肠排空时间延长。
此外,研究发现肌醇可以提高奥尼罗非鱼[6-7]、草鱼幼鱼[9]、异育银鲫[21]、幼建鲤[4]等的饲料效率。肌醇提高水生动物饲料效率的可能途径:1)通过提高消化吸收能力和蛋白质沉积率来促进饲料效率的提高[4];2)通过参与肝脏脂肪代谢来提高脂肪的利用率,从而提高饲料的利用率[6-8]。
1.2 肌醇缺乏与皮肤发育
饲料中肌醇缺乏通常还会引起许多水生动物出现皮肤出血、鳍条糜烂和体色发黑等症状[22]。这是因为:1)肌醇作为细胞膜的结构元素,通过酯化作用与细胞膜上的磷脂相结合形成肌醇磷脂酸盐,这种细胞膜元素的立体结构可能对维持细胞膜对离子和小分子的渗透能力起着重要作用。水生动物在缺乏肌醇时,体内的肌醇水平就会下降,使表皮的渗透能力降低,从而导致细胞破损。表现为皮肤出血糜烂。2)肌醇参与细胞膜之间的信号传递充当第二信使,当水生动物饲料中肌醇不足时,机体内的肌醇水平就会显著下降,从而影响细胞间的信号传递,机体自身的调节能力失衡[4]。
1.3 肌醇与脂肪代谢
营养性脂肪肝是现代养殖业中常见的疾病,在猪[23]、奶牛[24]、朗德鹅[25]、蛋鸡[26]、水貂[27]等动物均有报道。近几年,随着高密度水产养殖的迅速发展,养殖鱼类尤其是海水养殖鱼类,如鲈[28]、牙鲆[29]、大黄鱼[30]等的脂肪肝问题成为困扰养殖者的难题之一,鱼体脂肪肝降低了鱼肉品质和高温季节鱼类的抗应激能力,严重影响到鱼类养殖的效益。研究表明,饲料中营养素失衡和抗脂肪肝物质的缺乏是造成营养性脂肪肝的主要原因,营养不平衡的人工饲料(配合饲料或冰鲜小杂鱼虾)常常难以满足鱼体快速、健康生长的需要,造成养殖鱼类营养代谢紊乱。其中,肝脏脂肪代谢失调、沉积、浸润、脂肪含量升高,常常导致营养性脂肪肝发生[31-34]。
目前,有关鱼类营养性脂肪肝的研究主要围绕导致其产生的营养因素来进行,其中多数研究主要集中在宏量营养素,如蛋白质[29]、脂肪[35]、碳水化合物[36]等对鱼体脂肪代谢和体脂沉积的影响方面;其次是微量营养素与添加剂,如蛋氨酸、大豆卵磷脂[37]、溶血卵磷脂、甜菜碱和胆碱[38]等抗脂肪肝物质对鱼体脂肪沉积的改善作用方面。
肌醇为环已烷衍生物,具有很明显的亲脂性,能参与某些脂类代谢[39]。其参与肝脏脂质代谢的可能途径是肌醇使动物肝脏中的磷脂酰肌醇生成增多,然而,磷酸肌醇家族成员可作为脂质第二信使将效应器蛋白聚集于特定膜区以调控它们的组装或活性,它的增多导致极低密度脂蛋白的合成和分泌增多,而极低密度脂蛋白是肝脏脂肪转运的主要载体,使肝脏脂肪转运增强,肝脏脂肪代谢增强,反之则脂肪在肝脏积累,导致营养性脂肪肝[40]。
文华等[9]研究表明,饲料肌醇添加量≥200 mg/kg时草鱼幼鱼血清中总胆固醇和低密度脂蛋白固醇含量与对照组相比有显著提高,而血清甘油三酯含量与对照组相比有显著降低。吴宏玉等[8]对奥尼罗非鱼的研究发现,饲料中添加0.04%肌醇组肝脏中脂肪含量显著降低,未添加肌醇组的肝体比显著增加。崔红红等[41]研究表明,随着饲料中肌醇添加量的提高,团头鲂(Megalobrama amblycephala)肝脏中脂肪含量显著降低。
2 水生动物肌醇需要量及影响因素
从表2可以看出,杂食性和肉食性水生动物比草食性水生动物对肌醇的需要量高,可能是由于杂食性和肉食性水生动物的饵料中脂肪丰富,所以在长期进化中对参与脂肪代谢的肌醇需要量相对于草食性的水生动物高[3,5-17]。此外,目前研究显示水生动物对肌醇的需要量不仅随动物种类不同而不同,还受动物肌醇合成能力、生理状况、遗传上所造成的品系差异等的影响;一般以生长指标和肝脏肌醇蓄积量来确定水生动物对肌醇的需要量,某些研究也把水生动物血清指标[6,14]、消化酶和免疫酶活力[3-4,22]等作为肌醇需要量评定标准。
影响肌醇需要量的可能因素主要包括:1)肌醇的自体合成。大部分哺乳动物都具有肌醇合成能力并能满足其需要,而大部分水生动物依赖于饲料添加肌醇,这可能与它们缺乏肌醇合成能力有关。但经证实,如皱纹盘鲍[42]、斑点叉尾[5]自体合成的肌醇能满足其需要。2)肠道细菌的合成。在陆生动物中肠道细菌能提供合成肌醇给宿主动物[43]。3)饲料中的脂肪和糖类含量对肌醇需要量有一定影响[39]。

表2 水生动物肌醇的需要量Table 2 Myo-inositol requirements of aquatic animals
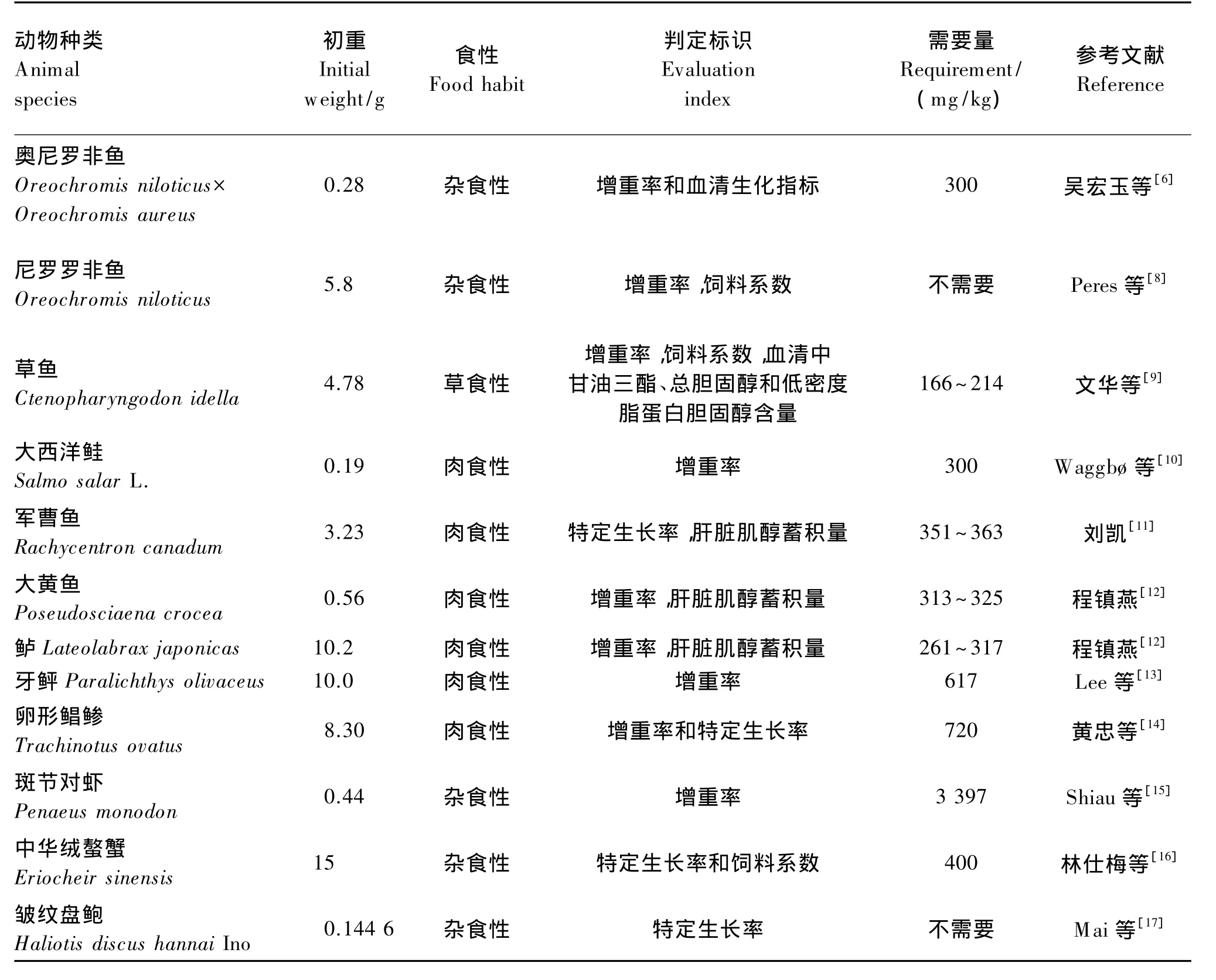
续表2
3 肌醇的来源与测定
大部分动植物组织中都含有肌醇。在植物组织中,肌醇主要以植酸的形式存在,植酸又名肌醇六磷酸,分子式为 C6H18O24P6,相对分子质量660.08[44]。植酸不能被有效利用,近来许多研究试图以植酸酶降解植酸,进而提高磷的利用率,并取得了一定效果,但添加植酸酶对肌醇利用率的影响尚未见报道。在动物组织中,肌醇主要以肌醇磷酸盐的形式存在,可以被有效利用[45]。
测定肌醇含量最经典使用的方法是重量法[46]和高碘酸钠氧化法[47],但这2种方法均较为繁琐,且不能排除邻二醇化合物如葡萄糖的干扰,往往使测定结果偏高。目前主要采取色谱法、酶法以及微生物法等测定肌醇含量,不仅简单、准确,重现性好,且能完全排除葡萄糖等杂质对肌醇测定的干扰。目前肌醇常用测定方法有:1)气相色谱法。试样用水在一定温度下振荡提取,过滤并定容。该提取液经烷基化衍生后,用气相色谱氢火焰检测器定性定量检测,最低检测限为2μg[48]。2)离子色谱法。肌醇分子具有电化学活泼型并可在强碱溶液中呈离子化状态,对其可采用高效阴离子交换色谱分离-脉冲安培检测(HPAEC-PAD)方法进行测定,最低检测限为1 ng[48]。3)高效液相色谱法。以 Lichrospher100 DIOL(150 mm×4.1 mm,5 μm,德国默克)为色谱柱,用蒸发光散射检测器(ELSD)进行测定,在10和25℃时的最低检测限分别为9和5 mg/L[49]。4)酶法。首先采用高氯酸等化学物质对试样进行前处理,然后利用肌醇脱氢酶与之反应,并在492 nm处采用分光光度计检测,该方法的测量范围为1~20 nmol[50]。5)微生物法。利用啤酒酵母(Saccharomyces cereviseae)对肌醇有特异需要的特性,测量范围为 20~200 mg/L[10]。
4 小 结
迄今,在水生动物中有关肌醇的研究主要还停留在需要量和对其生理功能影响的角度,而对其作用方式、病理机制和代谢途径等都缺乏比较系统和深入的探讨。目前,仅有Jiang等[51-58]通过体内和体外试验系统考察肌醇对幼建鲤肠道酶性抗氧化能力和非酶性抗氧化能力的影响及其影响幼建鲤肠上皮细胞抗氧化能力的作用方式。动物抗氧化系统包括酶性抗氧化系统和非酶性抗氧化系统,主要研究的酶性抗氧化系统包括超氧化物歧化酶(superoxide dismutase)、过氧化氢酶(catalase)、谷胱甘肽硫转移酶(glutathione-S-transferase)、谷胱甘肽过氧化物酶(glutathione peroxidase)和谷胱甘肽还原酶(glutathione reducase)等;非酶性抗氧化系统指还原性活性因子,如谷胱甘肽(glutathione)、维生素E(vitamin E)和维生素C(vitamin C)等。
然而,随着分子生物学技术的发展,从分子水平上阐明肌醇对主要水生动物代谢关键酶基因的多样性及其表达的调控机制,并从细胞水平阐述肌醇代谢的信号通路,对于在低等脊椎和无脊椎动物上微量营养物质的基础研究以及主要水生动物提高饲料效率和养殖效益具有重要意义。此外,通过比较发现,哺乳动物不需要肌醇,而绝大部分水产动物却依赖该种维生素。这些差异可能与它们生活环境的差异有关,涉及到渗透压调节等生理功能,这仍有待进一步研究。
[1] 刘晓永,钱和,刘建利.肌醇研究近况与展望[J].江苏食品与发酵,2003(4):7-10.
[2] NRC.Nutrient requirements of fish and shrimp[S].Washington,D.C.:National Academies Press,2011:206-207.
[3] 姜维丹.肌醇对幼建鲤消化吸收能力和免疫能力的影响[D].硕士学位论文.雅安:四川农业大学,2008.
[4] 李江.肌醇缺乏对幼建鲤消化功能和免疫功能的影响[D].硕士学位论文.雅安:四川农业大学,2007.
[5] BURTLE G J,LOVELL R T.Lack of response of channel catfish(Ictalurus punctatus)to dietary myoinositol[J].Canadian Journal of Fisheries and Aquatic Sciences,1989,46(2):218-222.
[6] 吴宏玉,唐瞻杨,杨鸿昆,等.肌醇对奥尼罗非鱼生长、肝脏和肌肉脂肪含量及血清生化指标的影响[J].南方农业学报,2011,42(11):1415-1419.
[7] SHIAU S Y,SU S L.Juvenile tilapia(Oreochromis niloticus×Oreochromis aureus)requires dietary myoinositol for maximal growth[J].Aquaculture,2005,243(1/2/3/4):273-277.
[8] PERES H,LIM C,KLESIUS P H.Growth,chemical composition and resistance to Streptococcus iniae challenge of juvenile Nile tilapia(Oreochromis niloticus)fed graded levels of dietary inositol[J].Aquaculture,2004,235(1/2/3/4):423-432.
[9] 文华,赵智勇,蒋明,等.草鱼幼鱼肌醇营养需要量的研究[J].中国水产科学,2007,4(5):794-800.
[10] WAAGBØ R,SANDNES K,LIE O,et al.Effects of inositol supplementation on growth,chemical composition and blood chemistry in Atlantic salmon,Salmo salar L.,fry[J].Aquaculture Nutrition,1998,4(1):53-59.
[11] 刘凯.军曹鱼(Rachycentron canadum)吡哆醇、肌醇和泛酸营养生理的研究[D].硕士学位论文.青岛:中国海洋大学,2008.
[12] 程镇燕.大黄鱼和鲈鱼对几种水溶性维生素营养需求及糖类营养生理的研究[D].博士学位论文.青岛:中国海洋大学,2010.
[13] LEE B J,LEE K J,LIM SJ,et al.Dietary myo-inositol requirement for Olive flounder,Paralichthys olivaceus(Temminch et Schlegel)[J].Aquaculture Research,2008,40(1):83-90.
[14] 黄忠,林黑着,牛津,等.肌醇对卵形鲳鲹生长、饲料利用和血液指标的影响[J].南方水产科学,2011,7(3):39-44.
[15] SHIAU S Y,SU S L.Dietary inositol requirement for juvenile grass shrimp,Penaeus monodon[J].Aquaculture,2004,241(1/2/3/4):1-8.
[16] 林仕梅,叶元土,罗莉,等.中华绒螯蟹对VC、VE、肌醇和胆碱需要量的研究[J].饲料工业,2000,21(8):21-23.
[17] MAI K S,WU G T,ZHU W.Abalone,Haliotis discus hannai Ino,can synthesize myo-inositol de novo to meet physiological needs[J].The Journal of Nutrition,2001,131(11):2898-2903.
[18] 陈四清,季文娟,吕用琦,等.肌醇对黑鲷幼鱼营养作用的研究[J].海洋科学,1999(5):13-15.
[19] 张美彦,王常安,徐奇友.肌醇对哲罗鲑生长性能、体成分及消化酶活性的影响[J].中国水产科学,2014(3):560-566.
[20] PEPPIATT C M,COLLINS T J,MACKENZIE L,et al.2-aminoethoxydiphenyl borate(2-APB)antagonises inositol 1,4,5-trisphosphate-induced calcium release,inhibits calcium pumps and has a use-dependent and slowly reversible action on store-operated calcium entry channels[J].Cell Calcium,2003,34(1):97-108.
[21] GONG W,LEI W,ZHU X,et al.Dietary myo-inositol requirement for juvenile gibel carp(Carassius auratus gibelio)[J].Aquaculture Nutrition,2014,20(5):514-519.
[22] 李江.肌醇缺乏对幼建鲤消化功能和免疫功能的影响[J].硕士学位论文.雅安:四川农业大学,2007.
[23] 班宏.中国猪肉行业质量管理现状与最优模式探讨[D].硕士学位论文.南京:南京农业大学,2006.
[24] 夏成,王哲,张洪友,等.患脂肪肝奶牛的代谢、内分泌和组织基因表达特征[J].中国农业科学,2010,43(8):1696-1702.
[25] 苏胜彦.朗德鹅肝脏脂平衡调节的初步研究[D].博士学位论文.南京:南京农业大学,2009.
[26] 刘健鹏,张勇.蛋鸡脂肪肝综合征的发病原因及预防措施[J].养禽与禽病防治,2013(4):39-41.
[27] 沙磊,徐永平,李淑英,等.饥饿和再投喂对水貂肝脏的影响研究[J].中国农学通报,2012,28(14):126-129.
[28] 杜震宇,刘永坚,田丽霞,等.饥饿对于鲈肌肉、肝脏和血清主要生化组成的影响[J].动物学报,2003,49(4):458-465.
[29] 邓君明.动植物蛋白源对牙鲆摄食、生长和蛋白质及脂肪代谢的影响[D].博士学位论文.青岛:中国海洋大学,2006.
[30] 曾端,麦康森,艾庆辉.脂肪肝病变大黄鱼肝脏脂肪酸组成、代谢酶活性及抗氧化能力的研究[J].中国海洋大学学报:自然科学版,2008,38(4):542-546.
[31] 王兴强,段青源,麦康森,等.养殖鱼类脂肪肝研究概况[J].海洋科学,2002(7):36-39.
[32] 吴强,彭西,崔恒敏.动物脂肪肝发病机理的研究进展[J].家禽科学,2006(4):46-48.
[33] 曾端.具有脂肪肝症状的几种海水养殖鱼脂类代谢和运输的研究[D].博士学位论文.青岛:中国海洋大学,2007.
[34] 黄春红,肖调义,胡毅,等.养殖鱼类脂肪肝研究现状分析[J].动物营养学报,2014,26(7):1715-1722.
[35] XU J H,QIN J,YAN B L,et al.Effects of dietary lipid levels on growth performance,feed utilization and fatty acid composition of juvenile Japanese seabass(Lateolabrax japonicus)reared in seawater[J].Aquaculture International,2011,19(1):79-89.
[36] 蒋利和,吴宏玉,黄凯,等.饲料糖水平对吉富罗非鱼幼鱼生长和肝代谢功能的影响[J].水产学报,2013,37(2):245-255.
[37] 曹俊明,林鼎,薛华,等.四种抗脂肪肝物质降低草鱼肝胰脏脂质积累的替代关系[J].水生生物学报,1999(2):102-111.
[38] 李红霞,刘文斌,李向飞,等.饲料中添加氯化胆碱、甜菜碱和溶血卵磷脂对异育银鲫生长、脂肪代谢和血液指标的影响[J].水产学报,2010,34(2):292-299.
[39] 李爱杰.水产动物营养与饲料学[M].北京:农业出版社,1996.
[40] 欧阳英石.磷酸肌醇家族与细胞功能[J].生物物理学报,2000,16(2):195-203.
[41] 崔红红,刘波,戈贤平,等.肌醇对团头鲂幼鱼生长、血清生化及组织成分含量的影响[J].上海海洋大学学报,2013,22(6):868-875.
[42] 吴格天.皱纹盘鲍(Haliotis discus hannai Ino.)水溶性维生素营养生理的基础研究[D].博士学位论文.青岛:青岛海洋大学,2001.
[43] NIELSEN E,BLACK A.Role of inositol in alopecia of rats fed sulfasuxidine[J].Experimental Biology and Medicine,1944,55(1):14-16.
[44] 吴澎,田纪春,王凤成.谷物中植酸及其应用的研究进展[J].中国粮油学报,2009,24(3):137-143.
[45] SHIEH T R,WARE J H.Survey of microorganisms for the production of extracellular phytase[J].Applied Microbiology,1968(9):1348-1351.
[46] 中华人民共和国药典委员会.中华人民共和国药典1977年版二部[M].北京:人民卫生出版社,1979.
[47] 胡利民,陈军.高碘酸法测定肌醇及其片剂的含量[J].中国药学杂志,1990,25(12):734-735.
[48] 中华人民共和国农业部.NY/T 1345—2007添加剂预混合饲料中肌醇的测定[S].北京:中国标准出版社,2007.
[49] PAZOUREK J.Fast separation and determination of free myo-inositol by hydrophilic liquid chromatography[J].Carbohydrate Research,2014,391:55-60.
[50] ASHIZAWA N,YOSHIDA M,AOTSUKA T.An enzymatic assay for myo-inositol in tissue samples[J].Journal of Biochemical and Biophysical Methods,2000,44(1/2):89-94.
[51] JIANG W D,FENG L,LIU Y,et al.Growth,digestive capacity and intestinal microflora of juvenile Jian carp(Cyprinus carpio var.Jian)fed graded levels of dietary inositol[J].Aquaculture Research,2009,40(8):955-962.
[52] JIANG W D,FENG L,LIU Y,et al.Myo-inositol prevents oxidative damage,inhibits oxygen radical generation and increases antioxidant enzyme activities of juvenile Jian carp(Cyprinus carpio var.Jian)[J].Aquaculture Research,2009,40(15):1770-1776.
[53] JIANG W D,FENG L,LIU Y,et al.Effects of graded levels of dietary myo-inositol on non-specific immune and specific immune parameters in juvenile Jian carp(Cyprinus carpio var.Jian)[J].Aquaculture Research,2010,41(10):1413-1420.
[54] JIANG W D,FENG L,LIU Y,et al.Lipid peroxidation,protein oxidant and antioxidant status of muscle,intestine and hepatopancreas for juvenile Jian carp(Cyprinus carpio var.Jian)fed graded levels of myoinositol[J].Food Chemistry,2010,120(3):692-697.
[55] JIANG W D,WU P,KUANG S,et al.Myo-inositol prevents copper-induced oxidative damage and changes in antioxidant capacity in various organs and the enterocytes of juvenile Jian carp(Cyprinus carpio var.Jian)[J].Aquatic Toxicology,2011,105(3/4):543-551.
[56] JIANG W D,LIU Y,JIANG J,et al.In vitro interceptive and reparative effects of myo-inositol against copper-induced oxidative damage and antioxidant system disturbance in primary cultured fish enterocytes[J].A-quatic Toxicology,2013,132/133:100-110.
[57] JIANG W D,KUANG S Y,LIU Y,et al.Effects of myo-inositol on proliferation,differentiation,oxidative status and antioxidant capacity of carp enterocytes in primary culture[J].Aquaculture Nutrition,2013,19(1):45-53.
[58] JIANG W D,LIU Y,HU K,et al.Copper exposure induces oxidative injury,disturbs the antioxidant system and changes the Nrf2/ARE(CuZnSOD)signaling in the fish brain:protective effects of myo-inositol[J].A-quatic Toxicology,2014,155:301-313.
