不同碳链长度n-3脂肪酸及脂肪水平对罗非鱼生长、肝功能和餐后血液指标的影响
2015-12-21李法见林仕梅彭祥和朱旺明
林 鑫 李法见 林仕梅* 彭祥和 朱旺明
(1.西南大学动物科技学院,重庆 400716;2.西南大学淡水鱼类资源与生殖发育教育部重点实验室,重庆 400716;3.广州市信豚水产技术有限公司,广州 510642)
研究表明,不饱和脂肪酸在机体内有着重要的生物学作用和营养生理调控功能,鱼油(fish oil,FO)与亚麻籽油(linseed oil,LO)是2种常见的饲料油脂,均富含n-3不饱和脂肪酸,但其碳链长度有所不同。鱼油中 C20∶5n-3(eicosapentaenoic acid,EPA)和 C22∶6n-3(docosahexaenoic acid,DHA)含量丰富。而亚麻籽油缺乏这2种脂肪酸,富含 α-亚麻酸(α-linolenic acid,ALA;C18∶3n-3)[1]。
不同碳链长度n-3脂肪酸会对鱼类产生不同的影响。已有研究表明,n-3脂肪酸对罗非鱼的最大生长是必需的[2]。目前,饲料中亚麻籽油可以完全替代鱼油而不影响养殖鱼类的生长[3-4],但也有研究发现亚麻籽油完全替代鱼油会对鱼类的生长产生不利的影响[5]。同鱼油相比,亚麻籽油会降低鲑鳟鱼肌肉脂肪含量[6],破坏肠道组织形态[7],但不会明显改变肝脏组织形态和韧性[8]。饲料脂肪的代谢主要依靠相关的酶来进行。n-3高不饱和脂肪酸(HUFA),如 EPA、DHA,均会抑制脂肪酸合成酶和葡萄糖-6-磷酸脱氢酶的活性[9],与含鱼油饲料相比,亚麻籽油能显著增加Δ6去饱和酶的活性,活化脂肪酸链延长通路,促进盲肠上皮细胞、肝细胞中C18∶2n-6和 C18∶3n-3生成C18∶3n-6和 C18∶4n-3[10]。众所周知,机体消化和吸收的脂肪及机体脂肪储备的动用都需经血液运输到其他组织,血清中脂肪存在的主要形式是胆固醇、甘油三酯(TG)、磷脂和自由脂肪酸等。因此,血脂水平可以反映机体脂肪代谢状况。目前有关饲料中添加亚麻籽油对鱼体餐后血液指标的影响未见报道。
为此,本试验以罗非鱼为试验对象,在实用饲料配方的基础上分别添加低水平(5%)、高水平(10%)的鱼油和亚麻籽油作为唯一脂肪源,比较不同碳链n-3脂肪酸及脂肪水平对罗非鱼生长、肝功能以及餐后血液指标的影响,以期从脂质代谢角度评价亚麻籽油对罗非鱼的作用效果。
1 材料与方法
1.1 试验饲料
以鱼粉、豆粕、棉籽粕和菜籽粕作为蛋白质源,配制基础饲料,在此基础上分别添加5%鱼油(低水平鱼油)、10%鱼油(高水平鱼油)、5%亚麻籽油(低水平亚麻籽油)和10%亚麻籽油(高水平亚麻籽油),配制成4种试验饲料。各饲料原料粉碎过40目筛,混合均匀,制成粒径为2.0 mm的硬颗粒饲料,风干后放入4℃冰箱中保存备用。试验饲料组成及营养水平见表1。

表1 试验饲料组成及营养水平Table 1 Composition and nutrient levels of experimental diets
1.2 试验鱼与饲养管理
试验鱼购自重庆北碚区鱼种场,训食适应环境10 d后,取体质健壮、规格整齐的罗非鱼[初始体重(43.70±1.06)g]360 尾,随机分成 4 组,每组设3个重复,每个重复30尾鱼。养殖试验在室内淡水循环玻璃水族缸(有效容积为300 L)中进行。日投饵量为鱼体重的4%~6%,每天08:30、12:30和16:30各投喂1次。养殖水源为曝气自来水。试验期间水温为(27.8±2.5)℃,pH 为 7.3±0.4,溶解氧浓度>6.3 mg/L,氨氮浓度<0.48 mg/L,亚硝酸盐氮浓度<0.06 mg/L。养殖周期为8周。
1.3 样品收集与分析
1.3.1 相关血液指标测定
8周饲养试验结束后,分别在最后一次投喂后3、6、9、12、15 和 24 h,从每个缸随机取 3 尾鱼(每组9尾鱼),称重并于尾静脉取血。采血时用浓度为100 mg/L的MS-222做快速深度麻醉,用一次性医用注射器从尾静脉采血。采出的血液直接采用安稳免调码血糖仪(三诺生物传感股份有限公司)对血糖(blood Glu)含量进行测定。剩余的血液制品置4℃冰箱过夜,于4℃条件下以6 000 r/min离心10 min,收集血清,-20℃保存备用。采用迈瑞BC-3000全自动生化分析仪检测血清中总胆固醇(TCHO)、TG、高密度脂蛋白胆固醇(HDL-C)、低密度脂蛋白胆固醇(LDL-C)以及极低密度脂蛋白胆固醇(VLDL-C)的含量。
1.3.2 常规生长指标测定
饲养试验结束后,禁食24 h,对剩下的鱼进行称重并计数,加上之前取样的鱼的体重,计算常规生长指标。
1.3.3 肝胰脏生化指标测定
称重后每缸随机取3尾鱼,取出肝胰脏,立即放入液氮罐中速冻,然后转入-80℃低温冰箱保存。肝胰脏匀浆液在2 500 r/min、4℃条件下离心10 min,取上清液作为酶活性分析样品,-20℃保存备用。肝胰脏谷丙转氨酶(ALT)、谷草转氨酶(AST)、超氧化物歧化酶(SOD)活性以及丙二醛(MDA)含量均采用南京建成生物工程研究所生产的试剂盒进行测定。组织中蛋白质含量采用考马斯亮蓝法测定。
1.4 计算公式
特定生长率(specific growth rate,SGR,%/d)=100×[ln末重(g)-ln初重(g)]/试验天数(d);
饲料系数(feed conversion ratio,FCR)=总干物质摄食量(g)/鱼体总增重(g);
蛋白质效率(protein efficiency rate,PER,%)=100×[末重(g)-初重(g)]/[总干物质摄食量(g)×蛋白质含量];
摄食量[feed intake,FI,g/(尾·d)]=总干物质摄食量(g)×2/[(鱼初始尾数+鱼终末尾数)×试验天数(d)];
100×总干物质摄食量(g)×2/{[末重(g)+初重(g)]×试验天数(d)};
成活率(survival ratio,SR,%)=100×终末鱼尾数(尾)/初始鱼尾数(尾)。
1.5 数据处理与分析
试验数据采用SPSS 17.0进行统计分析,结果以平均值±标准误(mean±SE)表示。脂肪源和脂肪水平对罗非鱼生长性能的影响采用两因素方差分析(two-way ANOVA)。脂肪源和脂肪水平以及取样时间对罗非鱼血液指标的影响采用三因素方差分析(three-way ANOVA),若有显著交互作用,则进行单因素方差分析(one-way ANOVA)。若差异达到显著水平,则进行Duncan氏法多重比较,显著性水平为 P<0.05。
2 结果与分析
2.1 不同碳链长度 n-3脂肪酸及脂肪水平对罗非鱼生长性能的影响
由表2可知,经过8周的饲养试验,各组罗非鱼的末重、SGR、FCR、PER均无显著差异(P>0.05)。脂肪水平显著影响罗非鱼的 FI(P<0.05),无论是鱼油还是亚麻籽油,其高水平组均显著低于低水平组(P<0.05)。试验期间,各试验组罗非鱼的成活率均为100%。
2.2 不同碳链长度 n-3脂肪酸及脂肪水平对罗非鱼餐后血液指标的影响
由表3可知,脂肪源、脂肪水平和取样时间对罗非鱼血清TCHO含量有极显著的影响(P<0.01),且脂肪源与脂肪水平间有显著的交互作用(P<0.05)。由图 1-a可知,对于血清 TCHO 含量,鱼油组高于亚麻籽油组,高水平组高于低水平组;在餐后24 h内,鱼油组峰值出现在餐后6 h,而亚麻籽油组则出现在餐后9 h。

表2 饲喂不同碳链长度n-3脂肪酸及脂肪水平饲料8周后罗非鱼的生长性能Table 2 Growth performance of tilapia fed the diets with different carbon chain length n-3 fatty acids and lipid levels after 8 weeks
由表3可知,脂肪源和取样时间对罗非鱼血清TG含量有极显著影响(P<0.01),且脂肪源与取样时间或脂肪水平间有极显著的交互作用(P<0.01)。由图1-b可知,对于血清TG含量,亚麻籽油组高于鱼油组;在餐后24 h内,各组峰值均出现在餐后6 h,随后呈下降趋势。
由表3可知,脂肪源、脂肪水平和取样时间对罗非鱼血清HDL-C含量均有极显著影响(P<0.01)。由图1-c可知,对于血清HDL-C含量,鱼油组高于亚麻籽油组,高水平组高于低水平组;在餐后24 h内,各组罗非鱼均在餐后6 h达到最低值。
由表3可知,脂肪源和脂肪水平对罗非鱼血清LDL-C含量分别有极显著(P<0.01)和显著影响(P<0.05),且脂肪源与脂肪水平间有显著的交互作用(P<0.05)。由图 1-d可知,对于血清LDL-C含量,鱼油组高于亚麻籽油组,高水平组高于低水平组。
由表3可知,脂肪源、脂肪水平和取样时间对罗非鱼血清VLDL-C含量分别有显著(P<0.05)、极显著(P<0.01)和极显著影响(P<0.01),且脂肪源与取样时间或脂肪水平间有极显著的交互作用(P<0.01)。由图 1-e可知,对于 VLDL-C 含量,亚麻籽油组高于鱼油组,高水平组高于低水平组;在餐后24 h内,各组VLDL-C含量均在餐后6 h达到峰值,随后亚麻籽油组迅速下降,而鱼油组在餐后9~15 h降低平缓。
由表3可知,脂肪水平和取样时间对罗非鱼血糖含量分别有显著(P<0.05)和极显著影响(P<0.01)。由图1-f可知,对于血糖含量,高水平组高于低水平组;在餐后24 h内,各组血糖含量随餐后时间的延长基本呈下降趋势。
2.3 不同碳链长度 n-3脂肪酸及脂肪水平对罗非鱼肝胰脏生化指标的影响
由表4可知,脂肪源极显著影响罗非鱼肝胰脏ALT活性(P<0.01),表现为鱼油组极显著低于亚麻籽油组(P<0.01);脂肪水平和取样时间对肝胰脏ALT活性没有显著影响(P>0.05)。各组罗非鱼肝胰脏AST活性无显著差异(P>0.05)。脂肪源、脂肪水平及取样时间均极显著影响罗非鱼肝胰脏MDA含量(P<0.01),且两两之间有极显著的交互作用(P<0.01),表现为高水平鱼油组>高水平亚麻籽油组>低水平鱼油组>低水平亚麻籽油组,组间差异极显著(P<0.01)。脂肪水平显著影响罗非鱼肝胰脏SOD活性(P<0.05),表现为低水平鱼油组显著高于高水平鱼油组(P<0.05),而高、低水平亚麻籽油组间差异不显著(P>0.05);脂肪源和取样时间对肝胰脏SOD活性没有显著影响(P>0.05)。

图1 饲喂不同碳链长度n-3脂肪酸及脂肪水平饲料罗非鱼餐后血液指标的变化Fig.1 Changes of postprandial blood indices of tilapia fed different carbon chain length n-3 fatty acids and lipid levels
3 讨论
3.1 不同碳链长度 n-3脂肪酸及脂肪水平对罗非鱼生长的影响
本试验结果表明,分别用低水平(5%)、高水平(10%)的鱼油或亚麻籽油饲喂罗非鱼8周,各组罗非鱼的生长性能没有显著差异。类似的研究结果在大西洋鲑[3]和石斑鱼[4]上已有报道。这可能是因为低水平鱼油饲料中已含有足够的HUFA,可满足罗非鱼正常生长的需求。另外,前人研究结果也证实了罗非鱼有能力使α-亚麻酸通过脱饱和及延伸途径转化为长链的HUFA(如DHA和EPA)[11],这可能是饲料中单独添加亚麻籽油时罗非鱼表现出与鱼油相同生长效果的原因。本试验中高水平鱼油或亚麻籽油并未表现出较好的促生长效果,这可能是因为高水平油脂会造成n-3与n-6脂肪酸比例失衡,超出罗非鱼对n-3脂肪酸的耐受限度,致使饲料中过多的脂肪被作为β-氧化的底物产能而消耗掉[12]。但也有研究发现,亚麻籽油完全替代鱼油会对鱼体生长产生不利的影响[5],这可能与试验饲料组成、鱼种以及养殖周期有关。
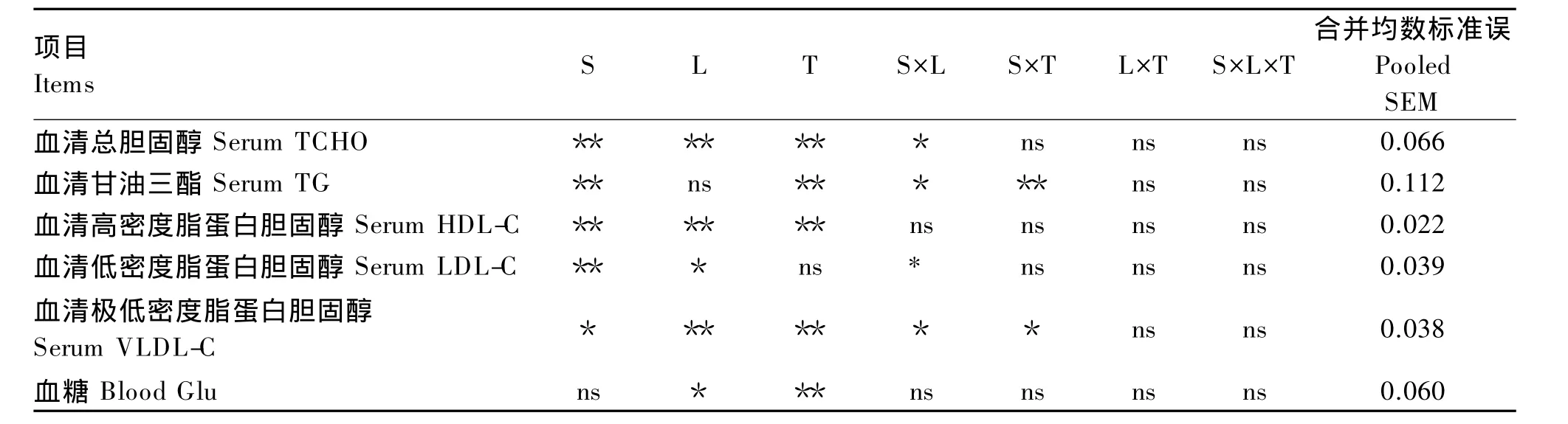
表3 脂肪源、脂肪水平、取样时间对罗非鱼餐后血液指标影响的方差分析Table 3 ANOVA of lipid source,lipid level and sampling time on postprandial blood indices of tilapia

表4 饲喂不同碳链长度n-3脂肪酸及脂肪水平饲料罗非鱼的肝胰脏生化指标Table 4 Hepatopancreas biochemical indices of tilapia fed the diets with different carbon chain length n-3 fatty acids and lipid levels
3.2 不同碳链长度 n-3脂肪酸及脂肪水平对罗非鱼餐后血液指标的影响
本试验结果表明,罗非鱼餐后血清TG含量明显升高,餐后6 h时达到峰值,而鲤鱼餐后8 h[13]、虹鳟餐后12 h血清TG含量才达到峰值[14]。TG峰值呈现时间差异可能与试验水温有关。本试验水温(28℃)较高,加快了罗非鱼脂质吸收速率[15],所以血清中TG含量达到峰值的时间先于其他的鱼类。硬骨鱼类肠道甘油三酯的合成和再酰化途径与哺乳动物类似[16]。一般脂肪酸的消化率会随着碳链长度的增加而降低,而亚麻籽油组罗非鱼血清TG含量高于鱼油组,这表明亚麻籽油中α-亚麻酸能够被肠道有效地吸收和再酰化[17]。本研究结果也表明,高水平亚麻籽油中的α-亚麻酸同样能被罗非鱼高效释放和有效利用,这与虹鳟[14]上的研究结果一致。
本试验结果发现,亚麻籽油组罗非鱼血清TCHO含量低于鱼油组,类似的结果在其他鱼类[18-20]上也被发现。这可能与植物油中胆固醇含量较低有关[21]。此外,鱼油组罗非鱼血清TCHO含量峰值出现的时间早于亚麻籽油组,这可能是因为鱼油组罗非鱼肝脏细胞摄取脂肪酸的能力强,致使β-氧化能力高于亚麻籽油组,这也是导致鱼油组罗非鱼血清TCHO含量高于亚麻籽组的另一个因素。众所周知,乙酰辅酶A是合成胆固醇的原料,而乙酰辅酶A也是脂肪酸β-氧化的产物。在大西洋鲑上的研究也发现,饲喂植物油的鱼体β-氧化能力显著低于鱼油[22]。进一步体外肝细胞试验证明,n-3高不饱和脂肪促进β-氧化是通过增强细胞脂肪酸摄取能力,而不是直接刺激β-氧化系统来完成的[23]。这些结果表明植物油替代鱼油会改变饲料或鱼体组织的脂肪酸组成,进而影响鱼体的β-氧化能力[21]。在哺乳动物上发现鱼油具有降低胆固醇作用,但在鱼类上还未见报道。研究发现,植物油能够降低美洲红点鲑肠道对胆固醇的吸收,进而导致血浆中和低密度脂蛋白(LDL)含量下降[19]。Panserat等[20]也发现,饲料以植物油为基础时,虹鳟肝脏胆固醇生物合成量较鱼油少。肝脏和肠黏膜是胆固醇合成的主要场所,体内胆固醇70%~80%由肝脏合成,10%由小肠合成,而血浆胆固醇含量反映了肝脏乙酰辅酶A的合成能力。有关饲料来源胆固醇或体内生物合成胆固醇的重要性还有待进一步研究。
在哺乳动物中,高密度脂蛋白(HDL)驱动内源性胆固醇从组织逆向肝脏转运,被称作“血管清道夫”。血浆HDL含量的高低与患心血管疾病的风险呈负相关。HDL同样也是硬骨鱼类血脂代谢的基础物质。本试验结果表明,鱼油组罗非鱼血清HDL-C含量高于亚麻籽油组,这与在尼罗罗非鱼[18]和虹鳟[14]上的研究结果类似。同椰汁油相比,鱼油或棕榈油也能够提高鼠血清中HDL-C含量[24]。此外,餐后6 h罗非鱼血清HDL-C含量达到最低值,可能是鱼油组餐后6 h血清TCHO含量达到峰值的原因之一,因为血清中缺乏HDL,不能及时驱动胆固醇逆转运,导致血清中TCHO含量升高。
LDL能够将胆固醇从肝脏转运到机体各组织中,而高水平的LDL-C会加重血管和组织的负担。本研究表明,鱼油组罗非鱼血清中不仅HDL-C含量高于亚麻籽油组,而且LDL-C含量也高于亚麻籽油组,这可能与亚麻籽油含有较高水平的α-亚麻酸有关。类似的结果在鲈鱼[25]和尼罗罗非鱼[18]上已有报道。在人类的研究上也发现,饱和脂肪酸(SFA)比不饱和脂肪酸(UFA)能够增加血清中TCHO和LDL-C含量[24]。而在猪上的研究表明,饲粮中补充不同脂肪源不会影响其血清中LDL-C含量[24]。血清中LDL-C含量与酰基辅酶A:胆固醇酰基转移酶(ACAT)有关,它是细胞内唯一合成胆固醇酯的酶,它优先选择UFA而不是SFA作为胆固醇酯化的底物[24]。因而,富含UFA的亚麻籽油饲料显著提高ACAT活性,促进胆固醇酯化形成LDL,进而增加了血清中LDL-C含量。另外,鱼油组血清LDL-C含量高于亚麻籽油组,可能还与鱼油组罗非鱼血清中TCHO含量高于亚麻籽油组有关。血液中90%的LDL是通过低密度脂蛋白受体(LDLR)途径清除的,而约50%的LDL在肝降解[24]。已有研究表明,饲料中脂肪酸也可通过调控LDLR介导来调节猪血浆中LDL含量,即SFA会降低肝脏LDLR活性,而多不饱和脂肪酸(PUFA)则会增强其活性[26]。鱼油和亚麻籽油是否也可以通过调控LDLR来调节罗非鱼血清中LDL含量,还有待研究。
研究表明,n-3 PUFA通过限制限速酶——甘油二酯酰基转移酶(DGAT)的活性以及肝脏中极低密度脂蛋白(VLDL)微粒的组装[27],进而抑制富含TG的VLDL微粒的分泌。然而,VLDL分解速率加快也可导致富含TG的脂蛋白含量减少[11]。本研究表明,鱼油组罗非鱼血清中不仅TG含量低于亚麻籽油组,而且VLDL-C含量也低于亚麻籽油组。鱼油富含DHA和EPA,它们能够通过刺激过氧化物酶体增殖物受体(PPAR)或固醇调节元件结合蛋白(SREBPs)通路抑制脂肪从头合成,进而促进脂肪酸的分解[28]。大西洋鲑上的离体细胞试验进一步证实,肝细胞释放脂肪的速率与饲料中脂肪源有关,鱼油组显著低于豆油组[29]。这可能是因为鱼油促使肝脏中脂肪酸的β-氧化作用增强,或者富含鱼油的饲料抑制VLDL中载脂蛋白B的合成,并促进周围组织的VLDL和肝脏的VLDL残体的清除[30]。本研究也进一步证实,餐后6 h各组罗非鱼血清中VLDL-C含量达到峰值,这主要是由于TG含量增高所致,同时还伴有HDL-C和血糖含量降低。
本研究表明,饲料脂肪水平能够显著影响餐后罗非鱼血糖含量。在虹鳟上的研究同样发现,饲料高脂肪水平通过提高肝脏葡萄糖-6-磷酸脱氢酶mRNA的表达量及其活性,进而显著增加血糖含量[31]。在哺乳动物中也发现,饲料中高脂肪水平会导致肝脏输出葡萄糖增加[32]。
3.3 不同碳链长度 n-3脂肪酸及脂肪水平对罗非鱼肝功能的影响
大量研究表明,肝胰脏转氨酶活性高低直接反映了肝胰脏受损伤的程度。本研究结果表明,亚麻籽油组罗非鱼肝胰脏ALT活性显著高于鱼油组,表明亚麻籽油能够改善罗非鱼肝功能,促进营养物质的消化利用,这与亚麻籽油富含α-亚麻酸密切相关。MDA是脂肪酸过氧化代谢产物,被认为是肝脏损伤的指标。本试验中,亚麻籽油组罗非鱼肝胰脏MDA含量显著低于鱼油组。这与在黑鲷上的研究结果[33]一致。此外,无论是鱼油还是亚麻籽油,高水平组罗非鱼肝胰脏MDA含量均显著高于低水平组。这表明高水平油脂(尤其是鱼油)在引起罗非鱼肝脏脂肪沉积的同时大量脂肪酸发生了过氧化。因鱼油富含DHA与EPA等HUFA,易受氧或其他自由基的攻击,产生对机体有害的过氧化产物,继而损害机体抗氧化能力[34]。通常鱼体内有天然的抗氧化系统去除这些氧化产物,以维持正常的生理功能,其中包括抗氧化酶SOD。动物细胞生存要求适当的氧化与抗氧化平衡,但脂肪酸被氧化的量一旦超过这种调节能力,肝脏就可能受到更严重的损伤。本试验结果显示,随饲料脂肪水平增加,鱼油组肝胰脏SOD活性显著降低。这表明鱼体处于氧化应激的状态,造成抗氧化能力下降[35]。相反,也有研究指出,HUFA能提高鲤鱼肝胰脏总抗氧化能力(T-AOC)[36],DHA 与 EPA 能够增强抗氧化酶活性,促进自由基清除[37]。到目前为止,有关DHA、EPA与机体抗氧化能力间的作用机制尚不清楚。
4 结论
饲料中不同水平的鱼油和亚麻籽油对罗非鱼的生长未产生负面影响,但会显著影响罗非鱼肝功能和餐后血液指标。
致谢:
感谢西南大学动物科技学院张小溪、杨振斌、张丹、苑鑫同学在饲料制备、养殖试验以及样品采集,杨阳、陈文燕同学在生化分析试验上的帮助。
[1] POPA V M,GRUIA A,RABA D N,et al.Fatty acids composition and oil characteristics of linseed(Linum usitatissimum L.)from Romania[J].Journal of Agroalimentary Processes and Technologies,2012,18(2):136-140.
[2] CHOU H Y,SHIAU SY.Both n-6 and n-3 fatty acids are required for maximal growth of juvenile hybrid tilapia[J].North American Journal of Aquaculture,1999,61(1):13-20.
[3] MENOYO D,LÓPEZ-BOTE C J,OBACH A,et al.Effect of dietary fish oil substitution with linseed oil on the performance,tissue fatty acid profile,metabolism,and oxidative stability of Atlantic salmon[J].Journal of Animal Science,2005,83(12):2853-2862.
[4] TURCHINI G M,FRANCIS D S,SENADHEERA S P S D,et al.Fish oil replacement with different vegetable oils in Murray cod:evidence of an“omega-3 sparing effect”by other dietary fatty acids[J].Aquaculture,2011,315(3/4):250-259.
[5] FRANCIS D S,TURCHINI G M,JONES P L,et al.Effects of dietary oil source on growth and fillet fatty acid composition of Murray cod,Maccullochella peelii peelii[J].Aquaculture,2006,253(1/2/3/4):547-556.
[6] TOCHER D R,BELL JG,DICK JR,et al.Polyunsaturated fatty acid metabolism in Atlantic salmon(Salmo salar)undergoing parr-smolt transformation and the effects of dietary linseed and rapeseed oils[J].Fish Physiology and Biochemistry,2000,23(1):59-73.
[7] OLSEN R E,MYKLEBUST R,KAINO T,et al.Lipid digestibility and ultrastructural changes in the enterocytes of Arctic char(Salvelinus alpinus L.)fed linseed oil and soybean lecithin[J].Fish Physiology and Biochemistry,1999,21(1):35-44.
[8] CABALLERO M J,OBACH A,ROSENLUND G,et al.Impact of different dietary lipid sources on growth,lipid digestibility,tissue fatty acid composition and histology of rainbow trout,Oncorhynchus mykiss[J].Aquaculture,2002,214(1/2/3/4):253-271.
[9] ALVAREZ M J,DIEZ A,LOPEZ-BOTE C,et al.Short-term modulation of lipogenesis by macronutrients in rainbow trout(Oncorhynchus mykiss)hepatocytes[J].British Journal of Nutrition,2000,84(5):619-628.
[10] OLSEN R E,RINGØ E.Lipids of arctic charr,Salvelinus alpinus(L.).Ⅱ.Influence of dietary fatty acids on the elongation and desaturation of linoleic and linolenic acid[J].Fish Physiology and Biochemistry,1992,9(5/6):393-399.
[11] LEAVER M J,BAUTISTA J M,BJÖRNSSON B T,et al.Towards fish lipid nutrigenomics:current state and prospects for fin-fish aquaculture[J].Reviews in Fisheries Science,2008,16(Suppl.1):73-94.
[12] BERGE G M,WITTEN P E,BAEVERFJORD G,et al.Diets with different n-6/n-3 fatty acid ratio in diets for juvenile Atlantic salmon,effects on growth,body composition,bone development and eicosanoid production[J].Aquaculture,2009,296(3/4):299-308.
[13] GEURDEN I,KAUSHIK S,CORRAZE G.Dietary phosphatidylcholine affects postprandial plasma levels and digestibility of lipid in common carp(Cyprinus carpio)[J].British Journal of Nutrition,2008,100(3):512-517.
[14] LUO L,XUE M,VACHOT C,et al.Dietary medium chain fatty acids from coconut oil have little effects on postprandial plasma metabolite profiles in rainbow trout(Oncorhynchus mykiss)[J].Aquaculture,2014,420/421:24-31.
[15] WALLAERT C,BABIN P J.Effects of temperature variations on dietary lipid absorption and plasma lipoprotein concentrations in trout (Oncorhynchus mykiss)[J].Comparative Biochemistry and Physiology Part B:Comparative Biochemistry,1994,109(2/3):473-487.
[16] CABALLERO M J,GALLARDO G,ROBAINA L,et al.Vegetable lipid sources in vitro biosyntheis of triacylglycerols and phospholipids in the intestine of sea bream(Sparus aurata)[J].British Journal of Nutrition,2006,95(3):448-454.
[17] OLSEN R E,HENDERSON R J,RINGØ E.The digestion and selective absorption of dietary fatty acids in Arctic charr,Salvelinus alpinus[J].Aquaculture Nutrition,1998,4(1):13-21.
[18] FERREIRA M W,DE ARAUJO F G,COSTA D V,et al.Influence of dietary oil sources on muscle composition and plasma lipoprotein concentrations in Nile tilapia,Oreochromis niloticus[J].Journal of the World Aquaculture Society,2011,42(1):24-33.
[19] GILMAN C I,LEUSCH F D L,BRECKENRIDGE W C,et al.Effects of a phytosterol mixture on male fish plasma lipoprotein fractions and testis P450scc activity[J].General and Comparative Endocrinology,2003,130(2):172-184.
[20] PANSERAT S,KOLDITZ C,RICHARD N,et al.Hepatic gene expression profiles in juvenile rainbow trout(Oncorhynchus mykiss)fed fishmeal or fish oil-free diets[J].British Journal of Nutrition,2008,100(5):953-967.
[21] TURCHINI G M,TORSTENSEN B E,NG W K.Fish oil replacement in finfish nutrition[J].Reviews in Aquaculture,2009,1(1):10-57.
[22] STUBHAUG I,LIE Ø,TORSTENSEN B E.Fatty acid productive value andβ-oxidation capacity in Atlantic salmon(Salmo salar L.)fed on different lipid sources along the whole growth period[J].Aquaculture Nutrition,2007,13(2):145-155.
[23] TORSTENSEN B E,STUBHAUG I.β-oxidation of 18:3n-3 in Atlantic salmon(Salmo salar L.)hepatocytes treated with different fatty acids[J].Lipids,2004,39(2):153-160.
[24] 张锐,张绍芬.低密度脂蛋白受体及其调节机制研究进展[J].国外医学:老年医学分册,2009,30(1):29-34.
[25] RICHARD N,KAUSHIK S,LARROQUET L,et al.Replacing dietary fish oil by vegetable oils has little effect on lipogenesis,lipid transport and tissue lipid uptake in rainbow trout(Oncorhynchus mykiss)[J].British Journal of Nutrition,2006,96(2):299-309.
[26] FERNANDEZ M L,WEST K L.Mechanisms by which dietary fatty acids modulate plasma lipids[J].The Journal of Nutrition,2005,135(9):2075-2078.
[27] KENDRICK J S,HIGGINSJA.Dietary fish oils inhibit early events in the assembly of very low density lipoproteins and target apoB for degradation within the rough endoplasmic reticulum of hamster hepatocytes[J].The Journal of Lipid Research,1999,40(3):504-514.
[28] JUMP D B,BOTOLIN D,WANG Y,et al.Docosahexaenoic acid(DHA)and hepatic gene transcription[J].Chemistry and Physics of Lipids,2008,153(1):3-13.
[29] VEGUSDAL A,GJØEN T,BERGE R K,et al.Effect of 18∶1n-9,20∶5n-3,and 22∶6n-3 on lipid accumulation and secretion by Atlantic salmon hepatocytes[J].Lipids,2005,40(5):477-486.
[30] 刘玉军,孙明堂,张枢泉,等.浓缩鱼油对高脂饲料大鼠血脂水平的影响[J].营养学报,1988,10(2):119-125.
[31] PANSERAT S,PERRIN A,KAUSHIK S.High dietary lipids induce liver glucose-6-phosphatase expression in rainbow trout(Oncorhynchus mykiss)[J].The Journal of Nutrition,2002,132(2):137-141.
[32] FANELLI C,CALDERONE S,EPIFANO L,et al.Demonstration of a critical role for free fatty acids in mediating counterregulatory stimulation of gluconeogenesis and suppression of glucose utilization in humans[J].Journal of Clinical Investigation,1993,92(4):1617-1622.
[33] PENG S M,CHEN L Q,QIN J G,et al.Effects of replacement of dietary fish oil by soybean oil on growth performance and liver biochemical composition in juvenile black seabream,Acanthopagrus schlegeli[J].Aquaculture,2008,276(1/2/3/4):154-161.
[34] SONG J H,FUJIMOTO K,MIYAZAWA T.Polyunsaturated(n-3)fatty acids susceptible to peroxidation are increased in plasma and tissue lipids of rats fed docosahexaenoic acid-containing oils[J].The Journal of Nutrition,2000,130(12):3028-3033.
[35] 吉红,周继术,曹福余,等.DHA对鲤抗氧化能力影响的初步研究[J].上海海洋大学学报,2009,18(2):142-149.
[36] 潘瑜,陈文燕,林仕梅,等.亚麻油替代鱼油对鲤鱼生长性能、肝胰脏脂质代谢及抗氧化能力的影响[J].动物营养学报,2014,26(2):420-426.
[37] JAHANGIRI A,LEIFERT W R,KIND K L,et al.Dietary fish oil alters cardiomyocyte Ca2+dynamics and antioxidant status[J].Free Radical Biology and Medicine,2006,40(9):1592-1602.
