产耐高温木聚糖酶菌株的筛选及其产酶条件优化
2013-04-23孙明哲郑宏臣孙君社裴海生刘逸寒路福平
孙明哲,郑宏臣,孙君社,3,裴海生,刘逸寒,张 璟,韩 杨,路福平*
(1.天津科技大学 生物工程学院 工业发酵微生物教育部重点实验室 工业酶国家工程实验室 天津市工业微生物重点实验室,天津 300457;2.农业部规划设计研究院,北京 100125;3.中国农业大学食品科学与营养工程学院,北京 100083)
木聚糖是被子植物(阔叶木和和本科)细胞壁中半纤维素的主要成分,其含量仅次于纤维素,是一种丰富的可再生资源[1]。作为一种异质性杂多糖,其主链是由D-木糖以β-1,4-糖苷键连接而成,其侧链由种类各异的糖基侧链组成[2]。内切-1,4-β-D-木聚糖酶作用于木聚糖主链,是木聚糖降解酶系中的关键酶,通常被简称为木聚糖酶[3]。木聚糖酶的来源相当广泛,已报道产木聚糖酶的微生物主要有细菌、放线菌、真菌、木霉、青霉、曲霉等,当前绝大多数商用木聚糖酶来源于霉菌,主要是由于霉菌微生物产酶量相对较高,但霉菌发酵周期长,并且大部分霉菌木聚糖酶最适作用条件偏于中性,使其在造纸、饲料、食品等工业的极端环境下应用受到一定限制[4-6]。而近年来报道的细菌木聚糖酶在耐极端条件方面有较强的优势,具有较好的开发前景[7]。
目前已报道的产木聚糖酶的细菌以芽孢杆菌居多。2012年,CHARBONNEAU DM等[8]在加拿大的肥料中筛选出多株产耐热木聚糖酶的细菌,其最适作用温度为60℃~65℃;2013年,ANAND A等[9]从印度的高温堆肥中筛选出一株嗜热脱氮芽孢杆菌(Geobacillus thermodenitrificansTSAA1),其所产木聚糖酶的最适温度为70℃。我国在产木聚糖酶细菌的研究方面也有显著成果,郑宏臣等[10]从造纸厂周边土壤中分离纯化到一株产碱性木聚糖酶的短小芽孢杆菌(Bacillus pumilusG1-3),并对其酶学性质进行分析,其最适反应pH值和温度分别为8.0和55℃;梁方方等[11]从长期堆放青贮饲料的土壤中分离到一株枯草芽孢杆菌X7,对其酶学性质的初步研究显示该酶最适pH值和温度分别为6.0和50℃;陈学敏等[12]从云南腾冲热泉水样中分离得到一株产木聚糖酶的菌株,并鉴定为地芽孢杆菌(Geobacillussp.PZH1),其木聚糖酶的最适pH值和最适温度分别为7.0和70℃;孙振涛等[13]从土壤中分离到一株产木聚糖酶的类芽孢杆菌(Paenibacillus campinasensis),其产木聚糖酶的最适pH值和最适温度分别为7.0和60℃。
本研究以筛选耐热木聚糖酶的产生菌为目的,从北京市顺义区的养牛场堆肥中获得一株可产耐热木聚糖酶的细菌,初步鉴定为类芽孢杆菌。对其木聚糖酶的酶学性质进行了初步研究,并优化了其发酵产酶的培养条件。同时本研究也进一步丰富了产木聚糖酶菌的菌种资源。
1 材料与方法
1.1 材料与试剂
牛粪样品:采集自北京市顺义区赵全营镇豹房村西北奶牛养殖场。
选择培养基:稻壳木聚糖1.00%,NH4NO30.50%,MgSO40.03%,NaCl 0.50%,K2HPO40.20%,(NH4)2SO40.10%,酵母粉0.03%,琼脂2.00%,pH值自然;种子培养基:牛肉粉0.50%,蛋白胨1.00%,NaCl 0.50%,pH值自然;基础产酶培养基:麸皮4.00%,蛋白胨1.00%,K2HPO40.50%,MgSO40.05%,NaCl 0.5%,pH值自然。
NaCl、NaOH、MgSO4、NH4NO3、K2HPO4、Na2HPO4、(NH4)2SO4、3-5-二硝基水杨酸(DNS)、苯酚、四水合酒石酸钾钠、偏重亚硫酸钠、尿素、柠檬酸均为国产分析纯;稻壳木聚糖购自天津市光复精细化工研究所;桦木木聚糖购自Sigma 公司;麸皮、玉米芯、玉米杆经粉碎后过40目筛。
1.2 仪器与设备
YT-CJ-2ND型洁净工作台、TU-1901双光束紫外可见分光光度计:北京普析通用仪器有限责任公司;HZQ-T160型恒温培养箱:哈尔滨东联电子技术开发有限公司法;BS 200 S型分析天平:北京赛多利斯天平有限公司。
1.3 方法
1.3.1 初筛方法
称1g牛粪样品于100mL无菌水中,振荡摇匀制得菌悬液。取1mL菌悬液加入到20mL灭菌的富集培养基中,于80℃水浴加热处理30min,以杀死不耐热的微生物营养细胞;然后置于50℃摇床上,培养2d。富集培养液经适当稀释后涂布于筛选平板培养基,置50℃恒温培养箱培养1d~2d,选取能产生透明圈的单菌落进一步划线纯化并编号,即木聚糖酶产生菌,作为进一步的复筛菌种。
1.3.2 复筛
将菌种活化16h后接入基础产酶培养基中进行摇瓶复筛,摇瓶使用100mL三角瓶装液50mL,180r/min、50℃,取72h发酵液4000r/min离心10min制得粗酶液,测其酶活力,选取酶活较高者保菌。
1.3.3 菌种鉴定
细胞形态观察参见微生物学试验技术[14]。16S rDNA序列分析:以产酶菌为模板,应用16S rDNA通用引物27F(AGAGTTTGATCMTGGCTCAG)和1492R(TACGGYTA CCTTGTTACGACTT),通过菌落PCR扩增产酶菌株的16S rDNA序列,PCR扩增程序:95℃、5min;94℃、45s,55℃、45s,72℃、90s,30个循环;72℃延伸10min。PCR结果送北京三博远志生物技术有限公司测序,测序结果在GenBank中进行比对。
1.3.4 Plackett-Burman试验
首先选择麸皮、玉米芯、玉米杆、稻壳木聚糖4种碳源及蛋白胨、牛肉膏、酵母浸粉、尿素、(NH4)2SO4、NH4NO36种氮源进行单因素试验,以确定最适的发酵产酶碳、氮源。
选取碳源浓度、氮源浓度、K2HPO4浓度、MgSO4浓度、NaCl浓度、温度、摇床转速、装液量、接种量、发酵初始pH值这10个因素进行Plackett-Burman试验[15](因素水平设计见表1)以确定影响发酵产酶重要因素。
1.3.5 响应面法优化发酵条件
根据Plackett-Burman试验的结果,确定影响发酵产酶的3个主要因素A、B、C,依据Box-Behnken中心组合设计原理[16],设计3因素3水平试验,共选取17个试验点,包含12个析因点,5个零点重复,用以估计试验误差。试验因素水平设计见表2。
1.3.6 木聚糖酶活力测定
木聚糖酶活力测定使用DNS法[7,17]。木聚糖酶活力单位的定义为:以1%的可溶性桦木木聚糖为底物,在酶的最适反应条件下,每分钟分解木聚糖生成1mol木糖所需酶量为一个酶活力单位(IU)。
2 结果与讨论
2.1 木聚糖酶产生菌的筛选
试验通过高温培养透明圈法从牛粪样品中筛选出木聚糖酶产生菌,其中选取28株产生透明圈较大的进行摇瓶发酵复筛,选取在高温条件下酶活相对最高的菌株NF1,并对其进行反复发酵验证后,发现菌株NF1在高温条件下产木聚糖酶较高且稳定性较好,该菌株发酵后制得的初酶液对桦木木聚糖底物的最适催化温度及pH值分别为60℃和6.0(见图1、图2),并且在高温条件下酶活可长时间保持稳定,说明菌株NF1可产耐高温木聚糖酶。
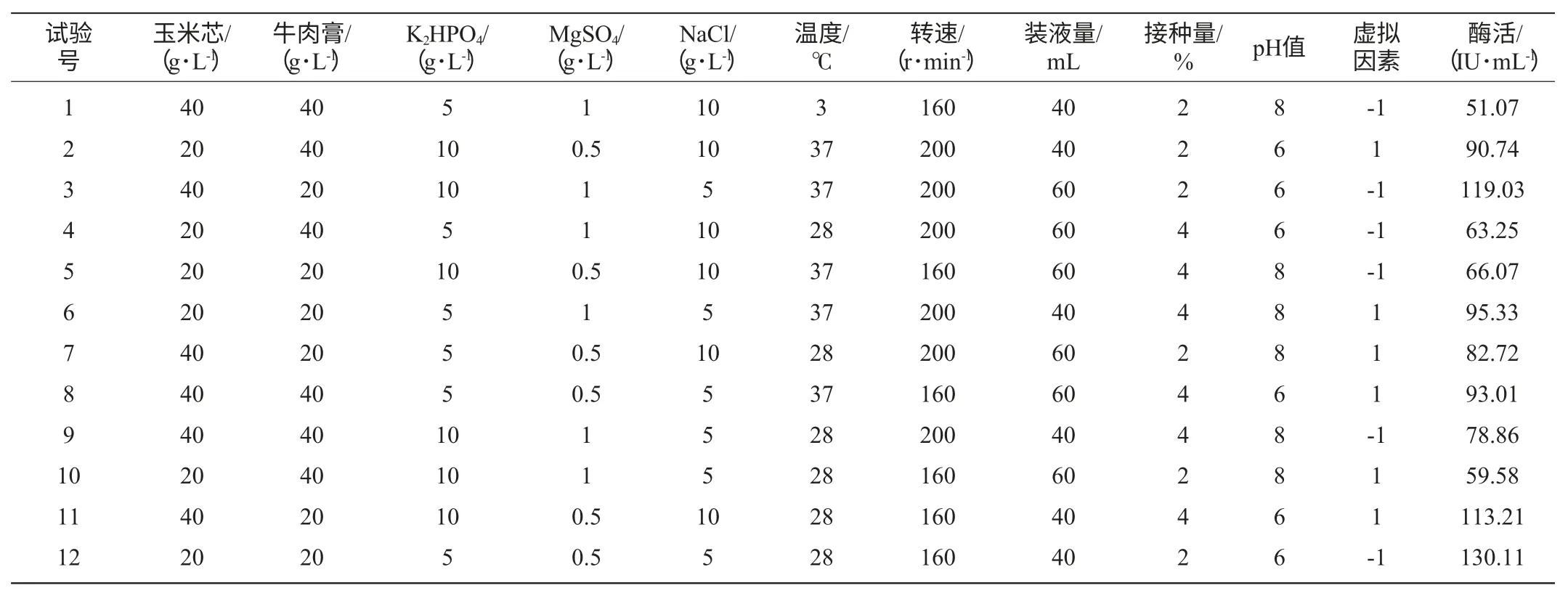
表1 Plackett-Burman试验设计及结果Table 1 Design and result of Plackett-Burman experiment
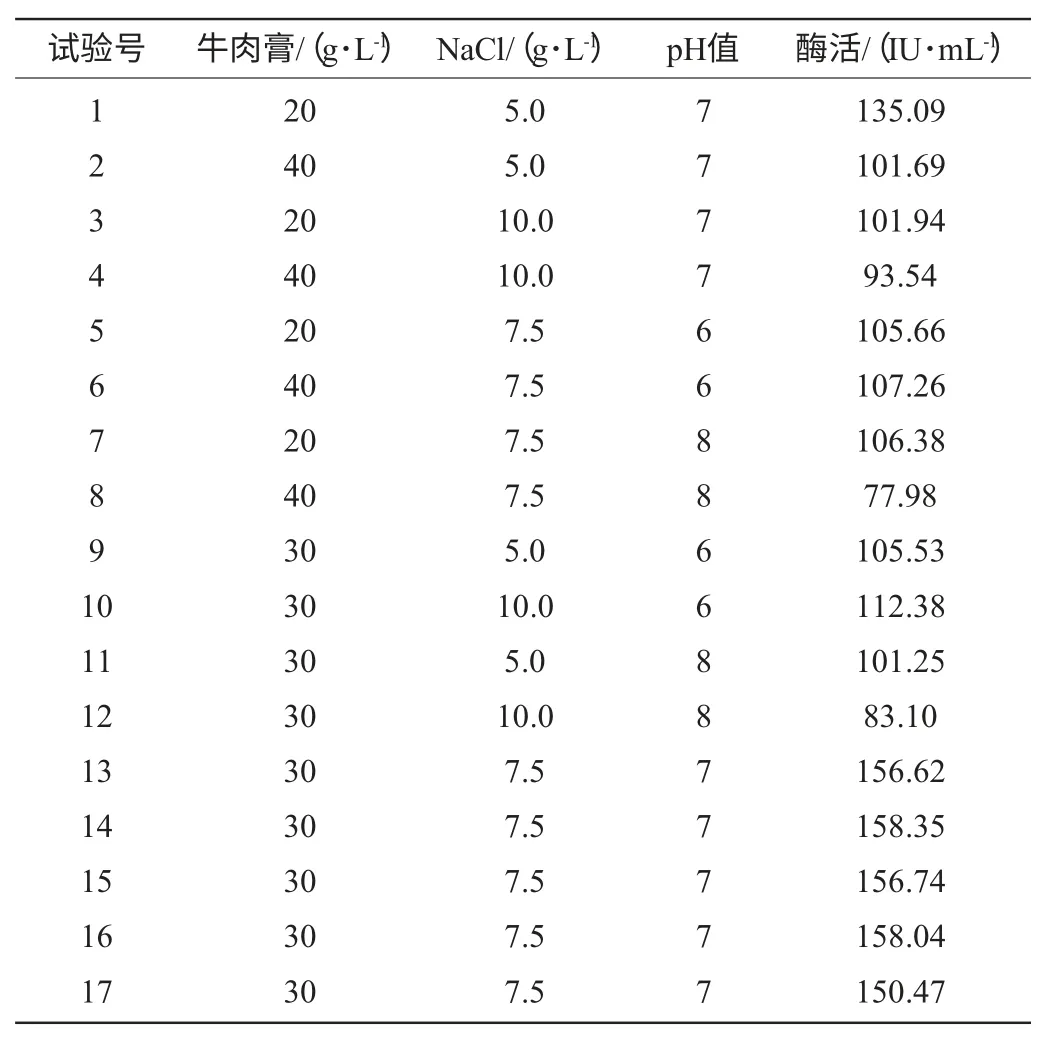
表2 Paenibacillus sp.NF1发酵产酶的Box-Behnken试验设计及结果Table 2 Experiment design and results of the Box-Behnken design for xylanase production
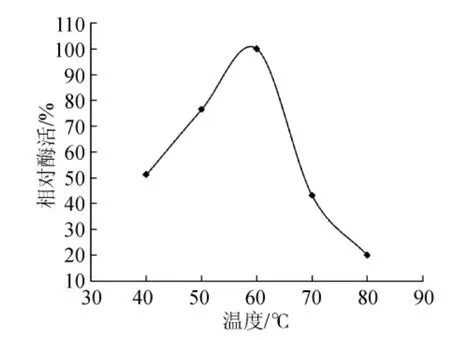
图1 最适催化温度Fig.1 The optimal catalytic temperature
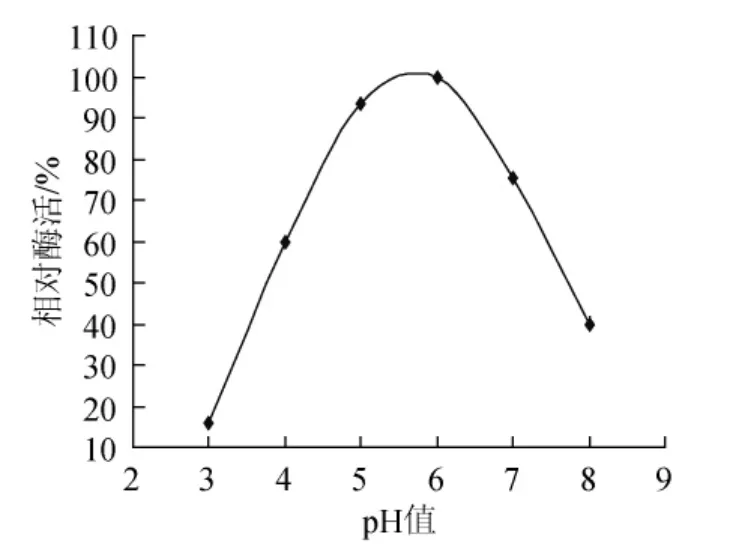
图2 最适催化pH值Fig.2 The optimal catalytic pH value
2.2 菌种鉴定
菌株NF1的菌体形态呈杆状,革兰氏染色阳性(图3);16S rDNA序列比对结果显示该菌属于类芽孢杆菌属,命名为类芽孢杆菌(Paenibacillussp.)NF1。

图3 菌株NF1的革兰氏染色图Fig.3 Gram’s staining of NF1
2.3 菌株NF1发酵产酶的Plackett-Burman试验
试验首先对Paenibacillussp.NF1发酵产酶的最佳碳、氮源进行了考察,结果见图4、图5。当使用稻壳木聚糖为碳源诱导产酶时发酵液酶活最高,但使用玉米芯为碳源时发酵液酶活与使用稻壳木聚糖相差不大,并明显高于使用麸皮和玉米杆的发酵液酶活,因此考虑到经济因素这里选择玉米芯为最佳碳源。氮源对发酵产酶的影响不是很大,选取酶活最高者牛肉膏为最佳氮源。确定最佳碳、氮源后,依据Plackett-Burman试验原理设计了试验,用以判定影响Paenibacillussp.NF1发酵产酶的主要因素,试验设计见表1。使用Design-Expert软件进行试验结果的分析,结果见表3。
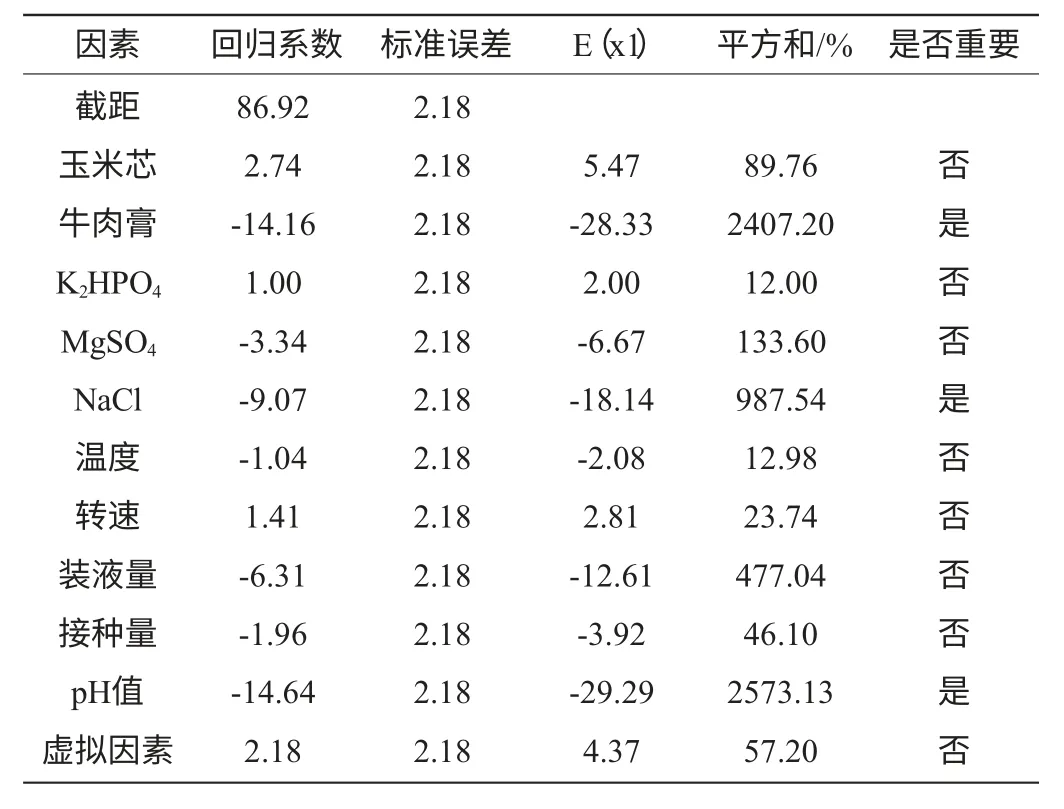
表3 偏回归系数及因素的显著性分析Table 3 Partial regression coefficients and analyses of their significance
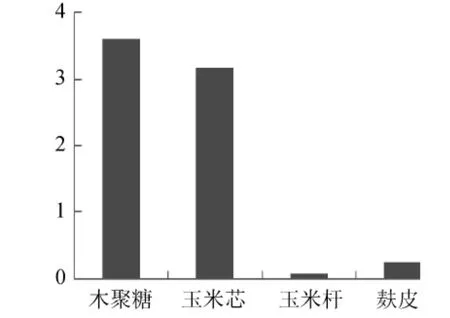
图4 不同碳源对Paenibacillus sp.NF1发酵产酶的影响Fig.4 Effect of different carbon source on Paenibacillus sp.NF1 xylnanse enzyme production
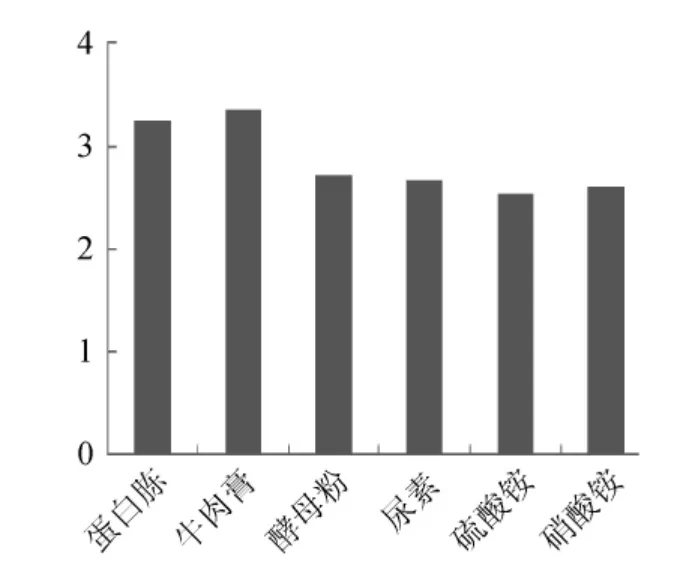
图5 不同氮源对Paenibacillus sp.NF1发酵产酶的影响Fig.5 Effect of different nitrogen source on Paenibacillus sp.NF1 xylnanse enzyme production and response values
以牛肉膏浓度因素为例,其偏回归系数为-14.16、标准误差为2.18、影响水平E(牛肉膏g/L)=-28.33,表明牛肉膏浓度对发酵产酶的影响为负效应,因此,在后续试验中应降低其水平;牛肉膏浓度的平方和百分比(contribution of SS)SS%=2407.20,明显高于玉米芯浓度、K2HPO4浓度、MgSO4浓度、温度、摇床转速、装液量、接种量等因素,因此显著性分析结果为重要。由表3可明显看出,牛肉膏浓度、NaCl浓度和发酵初始pH值为3个影响Paenibacillussp.NF1发酵产酶的主要因素,其影响值分别为2407.20%、987.54%、2573.13%,而虚拟因素对发酵产酶的影响值较低(57.20%)表明了该线性模型的适用性。
Plackett-Burman试验方差分析模型的Prob(P)>F值,为0.0006,表明所得回归方程达极显著水平,即该模型在被研究的整个回归区域拟合的很好;复相关系数(R2)=0.8750,说明相关性较好;校正相关系数(Adj R2)=0.8281,表明82.81%的试验数据的变异性可用此回归模型来解释;通常情况下变异系数(CV)越低,试验的可信度和精确度越高,CV=11.88%,表示PB试验的可信度和精确度较好;精密度(Adeq Precision)是有效信号与噪声的比值,大于4.0视为合理,本试验精密度为12.712(表4)。
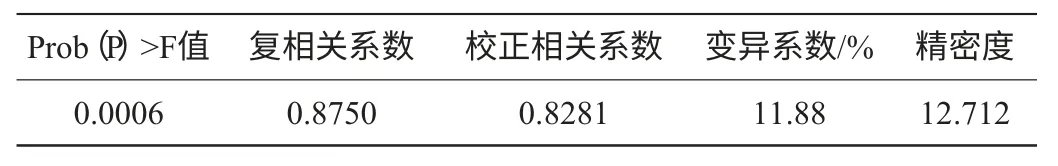
表4 Plackett-Burman试验方差分析Table 4 Analyses of variance for Plackett-Burman experiment
2.4 Paenibacillus sp.NF1发酵产酶条件的响应面分析
根据Plackett-Burman试验结果得出牛肉膏浓度、NaCl浓度、发酵初始pH值为影响Paenibacillussp.NF1发酵产酶的3个主要因素,按照Box-Behnken设计每个因素取3个水平进行试验,3因素3水平共17组试验,方案与结果见表2。
根据表2的试验结果,使用Design-Expert软件进行方差分析和二次回归拟合试验数据,回归方程变量分析见表5。
以木聚糖酶活力(R1)为响应面值,以A(牛肉膏浓度)、B(NaCl浓度)、C(发酵初始pH值)为自变量,拟合得到多元二次回归方程:R1=156.4-8.75A-6.57B-7.77C+6.25AB-7.50AC-6.25BC-24.61A2-23.36B2-32.11C2
由回归方程系数显著性检验及方差分析可知,模型试验拟合良好,模型的p<0.0001<0.05,表明该试验模型显著;失拟项p=0.0858>0.05,说明方程对试验的拟合度较好,此方法可靠。从表5中看到各因素对Paenibacillussp.NF1发酵产酶的影响不同,A、B、C的影响均为极显著;AB、AC、BC对发酵产酶的效应显著;A2、B2、C2也均对发酵产酶存在极显著影响。表明试验因子对响应值存在非线性关系,其中二次项与响应值也存在较大关系。模型的R2=0.9888,说明回归方程的拟合程度良好,失拟性较小。
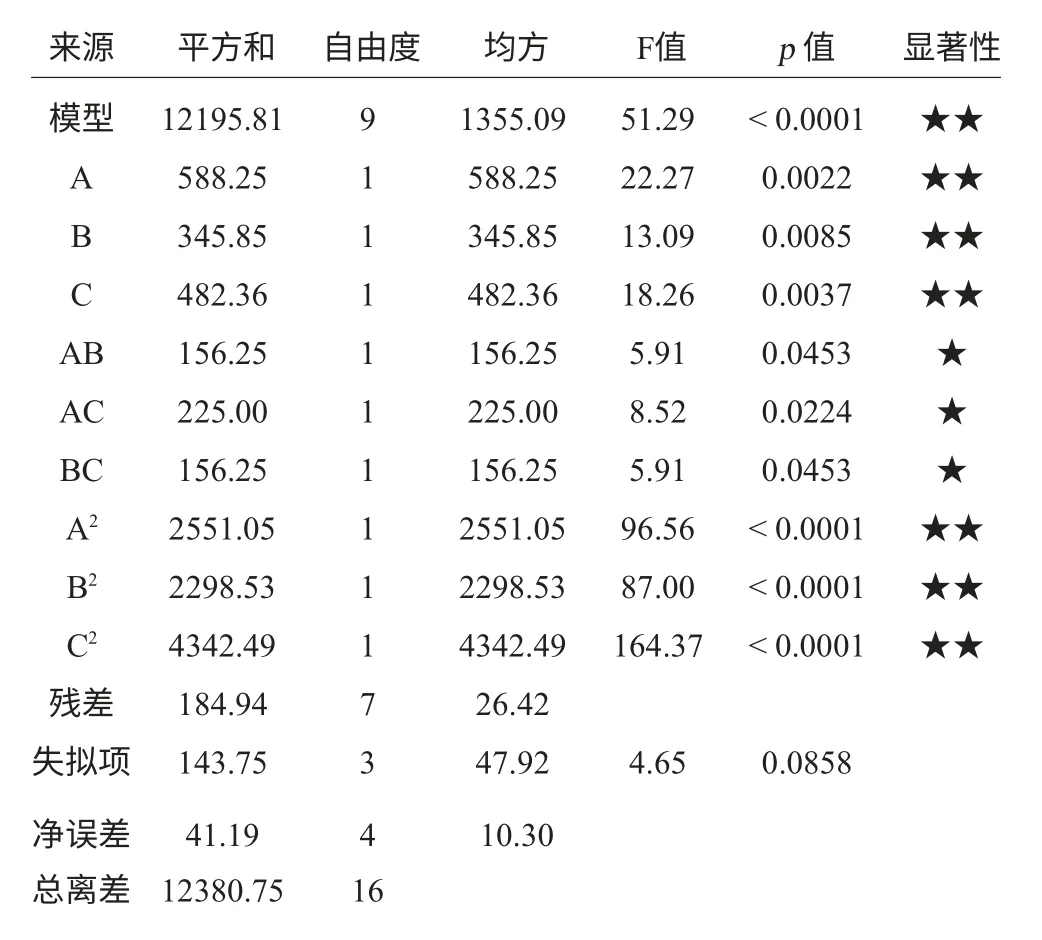
表5 回归方程变量分析Table 5 Variance analysis of regression equation
由图6可知,在发酵培养基初始pH值(pH=7.0)一定的条件下,木聚糖酶活力随NaCl浓度和牛肉膏浓度的升高而先升高后降低,两者交互作用显著,单因素效应相当。
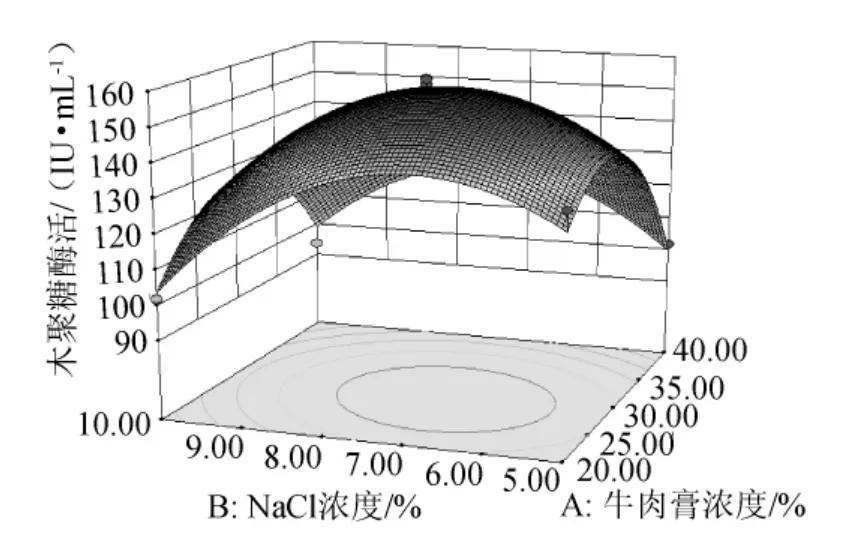
图6 牛肉膏浓度和NaCl浓度交互作用的响应面图Fig.6 Response surface map of the combined effects between concentration of beef extract and NaCl
由图7可知,在NaCl浓度一定的条件下,木聚糖酶活力随牛肉膏浓度和培养基初始pH值的升高而呈现先升高后降低的趋势,两者交互作用显著,且培养基初始pH值对木聚糖酶活力的影响比牛肉膏显著。
由图8可知,在牛肉膏浓度一定的条件下,木聚糖酶活力随NaCl浓度和培养基初始pH值的增加而呈想升高后降低的趋势,二者交互作用显著,单因素效应相当。
求解回归方程可得Paenibacillussp.NF1发酵产木聚糖酶的最优条件为牛肉膏28.19g/L,NaCl 7.12g/L,pH 值为6.92,此时模型预测的产酶最大值为157.653U/mL。对模型进行验证(取牛肉膏28.20g/L,NaCl 7.10g/L,pH值为6.90),设计3组平行,测得木聚糖酶酶活为160.6IU/mL,与预测酶活值偏差1.83%,无显著差异。经优化后Paenibacillussp.NF1的产酶量为原始培养条件的51.4倍。
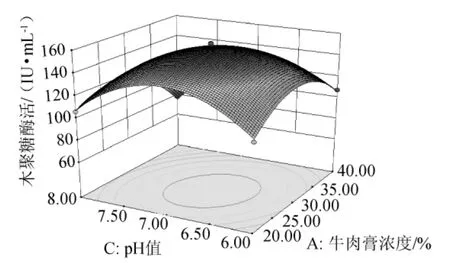
图7 牛肉膏浓度和培养基初始pH值交互作用的响应面图Fig.7 Response surface map of the combined effects between concentration of beef extract and initial pH value
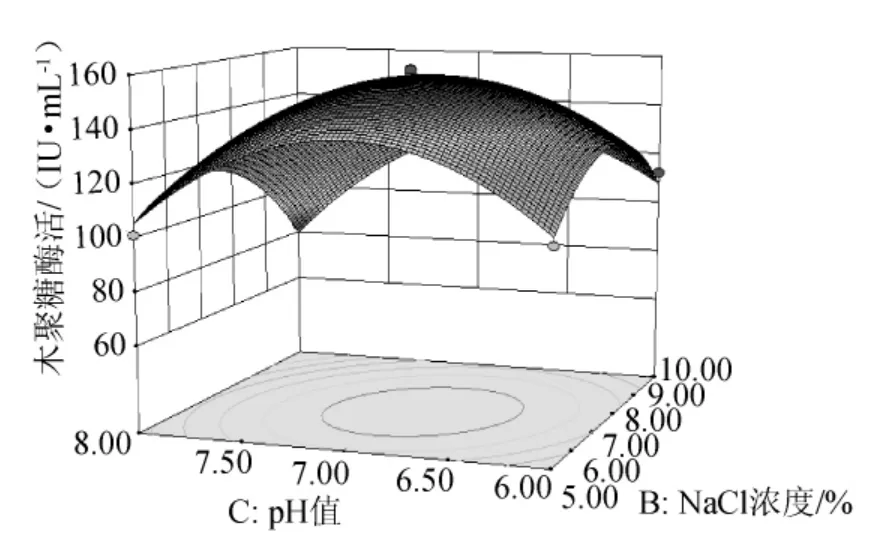
图8 NaCl浓度和培养基初始pH值交互作用的响应面图Fig.8 Response surface map of the combined effects between concentration of NaCl and initial pH value
3 结论
利用透明圈法从牛粪中筛选得到一株高产耐热木聚糖酶的菌株Paenibacillussp.NF1,其最适催化温度和pH值分别是60℃和6.0。应用响应面法对该菌发酵产木聚糖酶条件进行了优化,确定其最适发酵培养基组成为玉米芯20g/L、牛肉膏28.2g/L、K2HPO45g/L、MgSO40.5g/L、NaCl7.1g/L、培养基初始pH值为6.9;最适发酵条件为装液量100mL锥形瓶装液40mL、摇床转速160r/min、接种量2%、温度28℃。此条件下发酵72h,木聚糖酶活力可达160.6IU/mL,与预测结果一致,为原始发酵条件产酶量的51.4倍。
[1]GREGORY ACE,O’CONNELL AP,BOLWELL PG.Xylans[J].Biotechnol Genet Eng,1998,15(4):439-455.
[2]丁长河.链霉菌高产木聚糖酶及其酶学性质的研究[D].北京:中国农业大学博士论文,2003.
[3]BEG QK,KAPOOR M,MAHAJAN L,et al.Microbial xylanases and their industrial applications:a review[J].Appl Microbiol Biot,2001,56(3-4):326-338.
[4]SUNNA A,ANTRANIHIAN G.Xylanolytic enzymes from fungi and bacteria[J].Crit Rev Biotechnol,1997,17(1):39-67.
[5]SHI P,QIU Z,BAI Y,et al.A new xylanase fromStreptomyces megasporusDSM 41476 with high yield of xylobiose[J].World J Microb Biot,2012,28(2):687-692.
[6]MENON G,MODY K,KESHRI J,et al.Isolation,purification,and characterization of haloalkaline xylanase from a marineBacillus pumilusstrain,GESF-1[J].Biotechnol Bioproc Eng,2011,15(6):998-1005.
[7]ZHENG HC,LIU XG,HAN Y.Isolation,purification and characterization of a thermostable xylanase from a novel strainPaenibacillus campinasensisG1-1[J].J Microbiol Biotechn,2012,22(7):930-958.
[8]CHARBONNEAU DM,MEDDEB-MOUELHI F,BOISSINOT M,et al.Identification of thermophilic bacterial strains producing thermotoleranthydrolytic enzymes from manure compost[J].Indian J Microbiol,2012,52(1):41-47.
[9]ANAND A,KUMAR V,SATYANARAYANA T.Characteristics of thermostable endoxylanase and β-xylosidase of the extremely thermophilic bacteriumGeobacillus thermodenitrificansTSAA1 and its applicability in generating xylooligosaccharides and xylose from agroresidues[J].Extremophiles,2013,17(3):357-366.
[10]郑宏臣,刘逸寒,刘晓光,等.碱性木聚糖酶产生菌的筛选、XynG1-3 基因克隆表达及酶学性质研究[J].生物技术通报,2012(10):106-113.
[11]梁方方,莫 毅,杨 琳.木聚糖酶产生菌的筛选鉴定及酶学性质研究[J].饲料研究,2011(3):45-47.
[12]陈学敏,刘培培,张 波.一株产木聚糖酶嗜热菌的鉴定及酶学性质[J].微生物学通报,2011,38(2):151-156.
[13]孙振涛,刘建军,赵祥颖.一株产木聚糖酶菌株的分离、鉴定及其酶学特性研究[J].生物技术,2007,17(4):74-77.
[14]杜连祥,路福平.微生物学试验技术[M].北京:中国轻工业出版社,2008.
[15]孟 涛.关于Plackett-Burman 饱和设计数据分析方法的比较研究[D].上海:华东师范大学硕士论文,2008.
[16]黄新仁.响应面法在生物过程优化中的应用[D].长沙:湖南大学硕士论文,2011.
[17]BAILEY MJ,BIELY P,POUTANEN K.Interlaboratory testing of methods for assay of xylanase activity [J].J Biotechnol,1992,23(3):257-270.
