发酵温度与酶活性、酱油风味物质关系及其应用可能性
2012-04-13陶文沂倪海晴
陶文沂,倪海晴
(江南大学 生物工程学院,江苏 无锡 214122)
发酵温度与酶活性、酱油风味物质组成均有一定的内在关系。日本高盐稀态发酵的起始温度为10℃~15℃,发酵一定时间后调至20℃~25℃,最后升至30℃~35℃。产出的酱油酱香醇厚、酯香浓郁。由于发酵起始温度低,适宜低温环境的各种微生物和酶类都可能发挥作用,多种物质得到充分发酵,但是温度低导致发酵时间长,加大产品成本,与我国广大百姓消费水平不相适应。我国低盐固态发酵的品温在40℃~45℃,有利于提高原料利用率,周期短、成本低,但酱油香气的形成会受到一定的影响。笔者希望通过发酵温度与酶活性、酱油风味物质组成内在关系的探索,结合两者的工艺,合理调整发酵温度和盐水含量,改善我国低盐固态发酵香气不足的现状。
1 材料与方法
1.1 材料
菌种来源:米曲霉3.042:本实验室保藏菌种。
原材料:麸皮、豆粕、马铃薯均为市售,购于无锡市小三里桥粮油市场。
主要试剂:3,5-二硝基水杨酸,吐温80,L-酪氨酸来自国药集团化学试剂有限公司;乳酸来自中国医药集团上海化学试剂公司;无水碳酸钠来自上海虹光化工厂。
主要仪器与设备:SPX-150B-Z型生化培养箱,752型紫外分光光度计,超净无菌操作台,81-2型磁力恒温搅拌器,YXQ-LS-50SⅡ立式压力蒸气灭菌锅,迴转式恒温调速摇瓶柜等均为国产;液相色谱仪(Ag1100):美国安捷伦公司;高速离心机HITACHI CR22G:日本日立公司;光学显微镜:日本Olympus公司;2300自动定氮仪:FOSS-TECATOR。
培养基斜面培养基:马铃薯培养基(PDA);种曲培养基:麸皮20g,水19mL,拌匀后装于250mL三角瓶中,0.1MPa灭菌20min;大曲培养基:麸皮16g,豆粕24g,水32mL,拌匀后装于500mL三角瓶中,0.1MPa灭菌20min;酵母菌培养基:YPD培养基:葡萄糖20g/L,酵母提取物10g/L,蛋白胨20g/L。
主要试剂的配制。
(1)缓冲液:乳酸-乳酸钠缓冲液(pH 3.0):称取80%~90%乳酸10.6g,定容至1000mL为A液;称取70%乳酸钠16g,定容至1000mL为B液;取A液8mL、B液1mL混合,用蒸馏水稀释1倍,即成0.05mol/L乳酸-乳酸钠缓冲液。
磷酸盐缓冲液(pH 7.2):称取NaH2PO431.2g,定容至1000mL成0.2mol/LA液;称取Na2HPO471.63g,定容至1000mL成0.2mol/L B液;取A液28mL、B液72mL混合,用蒸馏水稀释1倍,即成0.1mol/L磷酸盐缓冲液(pH 7.2)。
醋酸钠缓冲液(pH 4.6):称取醋酸钠6.7g、冰醋酸2.6mL,蒸馏水溶解定容至1000mL。
上述缓冲液应以酸度计或精密试纸矫正pH值。
(2)2%酪蛋白溶液:中性酪蛋白溶液:称取干酪素2g,加入0.1 mol/L NaOH溶液10mL,小火加热煮沸使之溶解,用磷酸盐缓冲液(pH 7.2)定容至100mL。配置后应及时使用或冰箱保存。
2%酸性酪蛋白溶液:称取干酪素2g,加入1mL浓乳酸加速其溶解,然后加入少量乳酸-乳酸钠缓冲液(pH3.0),小火加热煮沸使之溶解,用乳酸-乳酸钠缓冲液(pH 3.0)定容至100mL,于4℃冰箱中保存备用。
(3)福林试剂:于2000 mL回流装置内,加入钨酸钠100g、钼酸钠25g、蒸馏水700mL、85%磷酸50mL、浓盐酸100mL,文火回流10h。加入硫酸锂50g,蒸馏水50mL,混匀后,加入几滴液溴,再煮沸15min驱逐残溴,并除去颜色,溶液应该呈黄色而不是绿色。若溶液仍有绿色,需再加几滴溴液,再煮沸除去之。待冷却后,定容至1000mL,过滤后置于棕色瓶中4℃冰箱保存,此溶液加2倍蒸馏水稀释即成稀释的福林试剂。
(4)DNS试剂:精确称取6.3g DNS(3,5-二硝基水杨酸)溶于262mL 2mol/L NaOH溶液中,然后加入酒石酸钾钠热溶液中(182.0g酒石酸钾钠溶于500mL水)。再加入5.0g苯酚和5.0g亚硫酸钠,加热搅拌至溶解,冷却后定容至1000mL,贮于棕色试剂瓶中4℃冰箱保存。
(5)1%可溶性淀粉:精确称取1.000g可溶性淀粉(以绝干计),加入醋酸钠缓冲液(pH 4.6),加热煮沸至透明,冷却后用醋酸钠缓冲液定容至100mL,此溶液需当天配制。
(6)0.4mol/L三氯乙酸(TCA)溶液:称取65.4g TCA,蒸馏水溶解定容至1000mL。
(7)6%苯酚:将苯酚加热溶解,冷却后准确称取6g苯酚,蒸馏水溶解定容至100mL。
(8)0.4mol/L碳酸钠溶液:称取无水碳酸钠42.4g,蒸馏水溶解定容至1000mL。
1.2 低盐固态发酵方法
1)制备孢子悬浮液:取一支试管斜面菌种,加入含0.1%吐温80作润湿剂的无菌生理盐水10mL,洗下孢子后倒入装有玻璃珠的100mL无菌三角瓶中,振荡器振荡30min后用砂芯漏斗过滤,调整孢子浓度为107个/mL。
2)制种曲:接种上述孢子悬浮液1mL于种曲培养基中,30℃培养72 h。
3)制大曲:大曲培养基灭菌后接入种曲,接种量为0.3%,搅拌均匀后,30℃培养48 h,即为发酵用大曲。
4)发酵:配制15%的盐水,按大曲:盐水=1:1.7加入盐水,搅拌均匀后于45℃水浴锅中发酵;48h后搅拌一次,然后加上保鲜膜密封发酵。
1.3 酶活的测定方法
1)含水量测定:取烘至恒重的培养皿,称取10g曲料后称得培养皿和曲料总重量为m1,105℃烘4.5h后称得总重量为m2,含水量
2)粗酶液制取方法:称取成曲5g,加入100mL蒸馏水,置于40℃水浴锅中浸提1h,间歇搅拌,用滤纸过滤得粗酶液。
3)蛋白酶测定方法:采用福林酚法。
蛋白酶活力单位定义:在40℃每分钟水解干酪素产生1μg酪氨酸的酶量,定义为1个蛋白酶活力单位。
蛋白酶活性测定:试管中加入已稀释的粗酶液样品1mL,于40℃水浴锅中预热2min,加入同样预热的酪蛋白溶液1mL,精确保温10min后立即加入0.4mol/L三氯乙酸2mL终止反应,40℃水浴中保温20min使残余蛋白质沉淀。离心,取上清液1mL加入0.4mol/L碳酸钠溶液5mL,福林试剂1mL,摇匀,40℃水浴保温发色20min,于波长660nm处测其吸光度值。空白试验方法同上,在加酪蛋白溶液之前先加0.4mol/L三氯乙酸2mL使酶失活后,再加入酪蛋白溶液。
测定酸性蛋白酶活性时,把稀释粗酶液和制备酪蛋白溶液用的缓冲液换成相应的缓冲液即可。中性蛋白酶测定时使用磷酸盐缓冲溶液(pH 7.2);酸性蛋白酶测定时使用乳酸-乳酸钠缓冲溶液(pH 3.0)。
蛋白酶活力计算公式:

式中:A为由粗酶液样品测得的OD值,查标准曲线得到相应的酪氨酸微克数;4 为4mL 反应液取出1mL 测定;n为粗酶液稀释倍数;10 为酶反应10min;ω 为样品中含水量。
在试验示范的跟踪过程中,云天化产品示范田的冬枣长势旺盛,中期表现出挂果多,果实品相好,树体健壮;采收期冬枣色泽好,果形均匀,畸形果少,口感佳。示范田亩增产82.4kg,亩增收10075.2元,给农户带来的一定的经济效益。通过试验示范,云天化“滴灌二铵”和“大量元素水溶肥”系列肥料得到了种植户的认可。
4)糖化酶测定方法采用DNS法测定。
糖化酶活力单位定义:在40℃每分钟水解可溶性淀粉产生1mg葡萄糖的酶量,定义为1个糖化酶活力单位。
样品测定:25mL比色管内加入1%可溶性淀粉溶液1.0mL,40℃水浴锅中预热10min,加稀释酶液1.0mL,40℃水浴中精确保温5min后,立即加入DNS溶液2.0mL终止反应,摇匀后沸水浴5min,冷却后用蒸馏水定容至25mL,摇匀后,在波长540nm处测其吸光度值。空白试验方法同上,在加酶液之前先加DNS溶液2.0mL以钝化酶活。
糖化酶活力计算公式为:

式中:A为由样品测得OD值,查标准曲线得相当的葡萄糖毫克数;n为酶液稀释的倍数;5 为反应5min;ω 为样品中水分含量。
1.4 游离氨基酸分析
取酱油样品1mL于25mL容量瓶中,用5%三氯乙酸溶液定容至25mL,沉淀2h后用双层滤纸过滤,于10000r/min离心10min后取上清液测定游离氨基酸含量。
分析方法OPA、FMOC柱前衍生化。
色谱条件 色谱柱:(250×4.6)mm,5μm,ODS HYPERSIL;柱温40℃;流速1.0mL/min;紫外检测器338nm,262nm(Pro,Hypro);洗脱方式:梯度洗脱。
1.5 酱醅分析
1)样品处理精确称取酱醅5.00g,放入150mL三角瓶中,加入70mL左右刚煮沸过的蒸馏水,浸泡0.5h,每隔10min搅拌一次。滤纸过滤,滤液移入100mL容量瓶中,加水定容,摇匀,即得5%滤液备用。
2)氨基酸态氮含量测定采用甲醛滴定法测定。
吸取上述5%滤液5mL放入250 mL烧杯中,加入蒸馏水60mL,用NaOH标准溶液电位滴定至pH值为8.2。然后,加入甲醛溶液10mL,混匀。再用NaOH标准溶液滴定至pH值为9.2,记下加入甲醛后消耗的NaOH标准溶液毫升数V2。空白试验方法同上,以5mL蒸馏水代替样品稀释液,记下空白试剂加入甲醛后消耗的NaOH标准溶液毫升数V1。
氨基酸态氮含量计算公式为:

式中:N为NaOH 标准溶液的当量浓度;W为吸取5%酱油样品的体积数,mL;0.014 为与1.00mL NaOH 标准溶液相当的氮的质量,g。
氢氧化钠标准溶液的标定:采用邻苯二甲酸氢钾标定法。
3)总酸含量测定:吸取上述5%滤液5mL放入250mL烧杯中,加入蒸馏水60mL,用NaOH标准溶液滴定至pH值为8.2。记下酱油样品滴定耗用NaOH标准溶液毫升数V2。空白试验方法同上,以5mL蒸馏水代替样品稀释液,记下空白试剂滴定耗用NaOH标准溶液毫升数V1。
总酸(以乳酸计)含量计算公式为:

式中:N为氢氧化钠标准溶液的当量浓度;W为吸取5%酱油样品的体积数,mL;0.09 为与1.00mL 氢氧化钠标准溶液相当的乳酸的质量,g。
4)总糖的测定:采用硫酸-苯酚法测定。将上述滤液稀释所需倍数,吸取2mL于比色管中,加入6%苯酚1.0mL和浓硫酸5.0mL。静置10min,摇匀后室温放置20min。然后于波长490nm处测其吸光度值。空白试验方法同上,以2mL蒸馏水代替样品稀释液。
5)还原糖的测定:采用DNS法测定。将上述滤液稀释所需倍数,吸取1mL于10mL比色管中,加入DNS试剂1mL,摇匀后置于沸水中精确保温5min。取出冷却后补加蒸馏水至10mL,摇匀后于波长540nm处测吸光度值。空白试验方法同上,以1mL蒸馏水代替样品稀释液。
6)总氮的测定:采用凯氏定氮法测定。
2 实验结果
2.1 三阶段温度条件下温度、盐水浓度与酶活性、酱油风味物质组成关系的初步探索
尝试采用发酵温度前期25℃、15d;中期30℃、10d;后期45℃、5d。制醅时添加盐水浓度为15%、21%、27%。同时以45℃、盐水浓度为15%的低盐固态发酵作为对照。跟踪测定发酵过程中蛋白酶、糖化酶、氨态氮含量、总酸、总糖、还原糖含量变化。
在发酵过程中,前10d每天取样,后20d每2d取一次样,测定蛋白酶、糖化酶活性的变化规律,结果见图1。

图1 发酵过程中中性蛋白酶活性变化曲线Fig.1 Neutral protease activity change in soy sauce fermentation
由图1可见,发酵前3d,尽管中性蛋白酶活力下降幅度都很大,但实验组酶活明显比对照组高,对照组第3d酶活82.26U/g干基,实验组酶活在500.00U/g~800.00U/g干基以上。20d时对照组检测不到酶活,而实验组在发酵结束时仍有酶活。
由图2可见酸性蛋白酶变化情况,对照组的酸性蛋白酶活力在发酵前3d明显下降,第7d就检测不到活性。实验组的酸性蛋白酶酶活在整个发酵过程中缓慢下降,其中27%盐浓度实验组第14d才没有酶活,其他2个实验组的酶活力延长至第20d。

图2 发酵过程中酸性蛋白酶活性变化曲线Fig.2 Acidic protease activity change in soy sauce fermentation

图3 发酵过程中糖化酶活性变化曲线Fig.3 Gluco-amylase activity change in soy sauce fermentation
由图3可见糖化酶变化情况,对照组的糖化酶活力在发酵前6d明显下降,实验组的酶活在前5d也显著下降,但下降幅度没有对照组大。随后,4组的糖化酶活力均缓慢下降,实验组的酶活力一直处于对照组的5倍以上。
由上可见,发酵温度对于酶活力有很大的影响。温度较低有利于酶活力的保存,对照组发酵温度偏高易造成酶失去活性。糖化酶最适作用温度较高,温度影响相对较小。
对应氨基酸态氮含量的变化可由图4分析。对照组的氨基酸态氮含量呈逐渐上升趋势;实验组在前期低温阶段氨基酸态氮含量增加有限。到第24d才由起始0.0333g/100mL增加为0.045g/100mL左右。在后期45℃发酵中增加较低温阶段明显。在整个发酵过程中,对照组的氨基酸态氮含量较实验组高出很多,最终相差约37%~63%。由此可见,低温虽然可以保持酶活较长时间,但低温不是酶作用最适温度,反应速率低使酶水解产物量也低。温度对于酶活、酶反应速率的关系是辩证的,很好地确定三者关系,有利于酱油发酵。
由图5、图6可见,总糖和还原糖在发酵过程中总体呈上升趋势,对照组的总糖和还原糖含量均较实验组高。20d时含量下降与发酵时美拉德反应和酒精发酵消耗有关。
发酵得到的酱油主要质量指标比较见表1。

图4 发酵过程中氨基态氮含量变化曲线Fig.4 Amino nitrogen content change in soy sauce fermentation

图5 发酵过程中总糖含量变化曲线Fig5 Total sugar content change in soy sauce fermentation

图6 发酵过程中还原糖含量变化曲线Fig.6 Reducing sugar content change in soy sauce fermentation

表1 实验组与对照组酱油质量指标比较Table 1 Comparison of indices of soy sauce between the experimental group and the blank group
由表1可知,实验组由于前期低温发酵,与对照组相比,总氮、氨基酸态氮含量等都有所下降。但是,实验组的氨基酸态氮生成率却高于对照组。可见:①低温发酵不利于酶反应进行,各项发酵产物含量均有所下降。②虽然实验组总氮含量不如对照组,原料利用率有所下降。但是实验组的氨基酸态氮生成率却高于对照组,显示,前期的低温发酵有利于使总氮转化为氨基酸态氮。
比较成品色泽外观可见,实验组酱油成品颜色较浅,体态澄清,浓度适中,酱香味轻微;而对照组酱油成品颜色较深,呈棕褐色,体态澄清,浓度大,酱香浓郁。
进一步对实验组与对照组酱油成品的游离氨基酸进行测定,结果及比较见表2。

表2 实验组与对照组低盐固态法发酵酱油游离氨基酸组成分析Table 2 Analysis of free amino acids composition in low-salt solidstate fermentation soy sauce between the experimental group and the control group
由表2可知,3组实验组的游离氨基酸总量都没有对照组高。但15%盐水含量的实验组的鲜味氨基酸含量为3.687mg/mL,较对照组3.223mg/mL高出14.39%,而苦味氨基酸较对照组降低35.23%;甜味和其他氨基酸也有所下降。其中,鲜味氨基酸的增加以及苦味氨基酸的减少可能是由于前期低温发酵的影响,低温发酵有利于适宜于低温环境的各种酶类充分发挥作用,因此,有利于各种氨基酸的浸出。
2.2 两阶段温度条件下温度与酶活性、酱油风味物质组成关系探索
由上面实验,前期25℃低温虽可长时间保存酶活,但低温条件下酶反应速率低,氨基酸态氮含量增长不大,30℃发酵阶段的氨基酸态氮含量开始有了较为明显的增加,所以,取消25℃,采用前期30℃发酵温度。后期45℃仅5d时间,氨基酸态氮含量明显不如低盐固态发酵的对照组,蛋白质一般经过8d~10d水解可基本完成,因此,适当延长后期45℃发酵时间为10d。在此基础上,第二次的探索实验分为2个阶段,发酵周期为30d,前期30℃、20d;后期45℃、10d。制醅同样采用15%、21%、27%3个盐浓度。跟踪测定发酵过程中蛋白酶、糖化酶、氨基酸态氮含量、总酸、总糖、还原糖含量的变化。
由图7~图9可见,第二次发酵过程酶活变化规律同第一次发酵相似,验证了低温对酶活的保护作用。实验组的中性蛋白酶活性在后期45℃的前2d有较大幅度下降;酸性蛋白酶酶活损失较快;在整个发酵过程中也都可以检测到糖化酶的活性。
总酸的含量变化见图10,在发酵过程中,总酸含量大体呈上升趋势,在个别测定点虽会有所下降,但下降程度不是很明显。对照组和3组实验组(15%、21%、27%)的总酸含量分别由起始的0.0415g/100mL、0.0241g/100mL、0.0291g/100mL、0.0291g/100mL增加到发酵结束时的0.1445g/100mL、0.0557g/100mL、0.0665g/100mL、0.0739g/100mL。

图7 发酵过程中中性蛋白酶活性变化曲线Fig.7 Neutral protease activity change in soy sauce fermentation
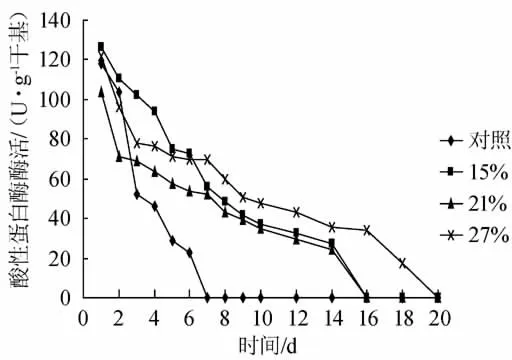
图8 发酵过程中酸性蛋白酶活性变化曲线Fig.8 Acidic protease activity change in soy sauce fermentation

图9 发酵过程中糖化酶活性变化曲线Fig.9 Glucoamylase activity change in soy sauce fermentation

图10 发酵过程中总酸含量变化曲线Fig.10 Total acid content change in soy sauce fermentation
由图11所示,第二次发酵过程中氨基酸态氮含量变化的规律较第一次明显。整个过程中氨基酸态氮含量呈上升趋势。对照组始终高于实验组,说明传统的45℃发酵有利于氨基态氮的生成。对照组的氨基酸态氮含量由起始的0.0427g/100mL上升到最终的0.0861g/100mL;15%、21%、27%盐水含量的实验组分别上升到0.0684g/100mL、0.0657g/100mL、0.0508g/100mL。3组实验组在发酵过程中的对比,氨基酸态氮含量的高低大体为15%盐浓度最高,说明高的食盐含量降低了酶反应速率,同时也抑制产物氨基酸态氮的生成。在以后的实验中,采用15%的盐浓度。

图12 发酵过程中总糖含量变化曲线Fig.12 Total sugar content change in soy sauce fermentation

图13 发酵过程中还原糖含量变化曲线Fig.13 Reducing sugar content change in soy sauce fermentation
图12、图13所示规律与第一次实验相同。实验组的还原糖和总糖含量在18d左右,对照组在22d左右有所下降。也是由于美拉德反应和酵母酒精发酵造成的。
将实验组与45℃发酵对照组进行低盐固态发酵,得到酱油主要质量指标比较见表3。这批实验组延长了后期45℃发酵时间,酱油成品的色泽、酱香味与对照组不相上下。
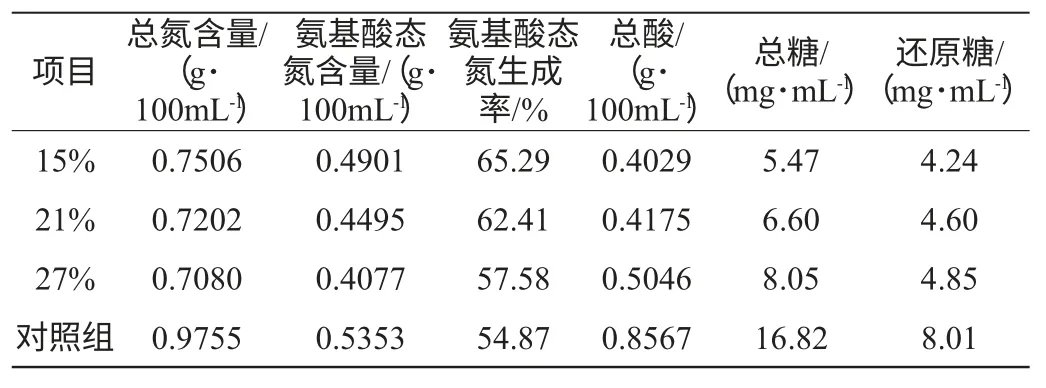
表3 实验组与对照组低盐固态法发酵酱油质量指标比较Table 3 Comparison of indices of low-salt solid-state fermentation soy sauce between the experimental group and the blank group
由表3可知,这批发酵的氨基酸态氮、总氮和总酸含量较第一次实验效果明显提高,说明温度变化设计较上次合理。其中,15%盐水含量的实验组的氨基酸态氮含量与对照组的最为接近。同第一次实验一样,实验组的氨基酸态氮生成率均比对照组的高,说明了低温可能更好地促进总氮转化为氨基酸态氮。4组酱油成品游离氨基酸分析结果见表4。

表4 实验组与对照组低盐固态法发酵酱油游离氨基酸组成分析Table 4 Analysis of free amino acids composition in low-salt solidstate fermentation soy sauce between the experimental group and the control group
由表4可知,改进后的实验较第一次实验的游离氨基酸总量明显提高。虽仍低于对照组,但值得关注的是,鲜味氨基酸的含量较对照组有大幅度提高,以15%盐水含量的实验组效果最为明显,其鲜味氨基酸含量7.949mg/mL,较对照组的6.546mg/mL提高了21.42%;苦味氨基酸、甜味氨基酸及其他都有所减少。另外,21%盐水含量的实验组的鲜味氨基酸也比对照组的高。21%、27%盐水含量的实验组的甜味氨基酸、其他氨基酸有所减少,苦味氨基酸有所增加。由于谷氨酸赋予了酱油的鲜味尤为重要,因此综合考虑,15%盐水含量的实验组最为理想,在下一步的工作中采用15%的盐水含量。
2.3 改进的两阶段温度条件下温度与酶活性、酱油风味物质组成关系
在前两次实验的基础上,对方案进行进一步调整。采用15%的盐水浓度,将45℃发酵时间进一步延长为15d;发酵分为2个阶段,30℃发酵和45℃发酵。实验方案:
①前期30℃发酵,时间分别为5d、10d 2个实验组,后期都为45℃发酵15d,发酵周期分别为20d、25d,目的是考察前期低温发酵酶活保持稳定对于发酵的影响。
②前期都为45℃发酵15d,后期低温30℃发酵,时间分别为5d、10d 2个实验组,发酵周期分别为20d、25d,目的是考察前期低温和后期低温对于酵母菌、乳酸菌繁殖的效果。
同时以45℃低盐固态发酵为对照实验。
按照上述实验方案进行低盐固态发酵,得到的酱油主要理化指标比较见表5。

表5 不同发酵周期酱油质量指标比较Table 5 Comparison of indices of different fermentation periodic soy sauce
由表5可知,前高温后低温的实验组较前低温后高温的实验组各项指标略高;25d发酵的各项理化指标比20d发酵有所提高,且与对照组的更为接近。其中,前高温后低温发酵酱油成品较前低温后高温的总糖含量、还原糖含量高出很多,且前高温后低温、25d发酵酱油成品的总糖、还原糖含量较对照组的高1.36mg/mL、2.83mg/mL,这将有利于后期酵母酒精发酵及产醇、产酯。因此,采用前高温后低温、25d的发酵方式较为合理。
按照上述实验方案进行低盐固态发酵,得到的酱油游离氨基酸分析见表6。

表6 不同发酵周期酱油游离氨基酸组成分析Table 6 Analysis of free amino acids composition in different fermentation periodic soy sauce
由表6可知,25d发酵酱油的游离氨基酸总量比20d发酵酱油有所提高。这说明,随着发酵时间的延长,有利于游离氨基酸的生成。虽然前低温后高温的发酵中鲜味氨基酸的增加、苦味氨基酸的减少较前高温后低温的效果明显,但后者与对照组相比,效果也很明显。结合2种发酵方式的酱油理化指标比较,确定可参考的新工艺参数为:前期45℃发酵15d,后期30℃发酵10d,发酵周期为25d。
3 发酵温度与酶活性、酱油风味物质组成内在关系研究的应用可能性
由发酵温度的初步探索实验结果可知,前期低温对于蛋白酶活性的保持有很明显的效果。而对照组整个发酵过程保持45℃,不利于蛋白酶活性的保持,但是低温会降低酶促反应的速率,因此实验组发酵过程中和酱油成品的理化指标较对照组有所下降。其中,氨基酸态氮在前期的低温阶段氨基态氮含量增加有限,在后期45℃发酵中,氨基酸态氮含量的增加较低温阶段明显。对于酱油成品,虽然实验组的总氮含量不如对照组,但氨基酸态氮生成率却高于对照组,所以,前期低温发酵能很好地使总氮转化为氨基酸态氮。由游离氨基酸分析可知,3组实验组的游离氨基酸总量都没有对照组高。但15%盐水含量实验组的鲜味氨基酸含量为3.687mg/mL,较对照组3.223mg/mL高出14.39%,苦味氨基酸较对照组降低了35.23%;甜味和其他氨基酸也有所下降。由于前期25℃发酵阶段氨基酸态氮含量增长不多,30℃发酵阶段氨基酸态氮含量开始有了较为明显的增加。因此,可以摒弃25℃发酵阶段,前期采用30℃发酵,后期45℃发酵即可。
由两阶段发酵温度探索实验可知,由于温度的调整,发酵过程以及酱油成品的各项指标均优于第一次实验。其中,15%盐水含量实验组的鲜味氨基酸含量为7.949mg/mL,较对照组的6.546mg/mL提高了21.42%;苦味氨基酸含量为5.574mg/mL,较对照组的6.977mg/mL减少了20.12%。
由改进的两阶段发酵温度实验可知,前高温后低温、25d发酵酱油较对照组相比,鲜味氨基酸增加了21.58%,苦味氨基酸减少了21.61%,总糖含量、还原糖含量也比对照组含量高。综合考虑各种游离氨基酸含量及酱油各种理化指标,确定采用前期45℃发酵15d,后期30℃发酵10d,发酵周期25d的新工艺是有可能性的。
(略)
