乳酸菌对发酵乳制品风味形成的影响
2012-01-08李良马莺
李良,马莺
(1.哈尔滨工业大学食品科学与工程学院,哈尔滨 150090;2.东北农业大学食品学院,哈尔滨 150030)
乳酸菌对发酵乳制品风味形成的影响
李良1,2,马莺1
(1.哈尔滨工业大学食品科学与工程学院,哈尔滨 150090;2.东北农业大学食品学院,哈尔滨 150030)
乳酸菌对发酵乳制品风味有决定性影响,发酵乳制品的风味物质主要是乳酸菌利用脂肪、碳水化合物和蛋白质等物质产生的。文章对乳酸菌代谢脂肪、糖、柠檬酸和氨基酸产生芳香物质的机理进行了综述。
乳酸菌;风味;机理
0 引言
发酵乳制品是以鲜乳或还原乳为主要原料,添加适当蔗糖等配料后经乳酸菌发酵制成的乳制品[1],包括酸乳、酸奶油和酸奶等产品。发酵乳制品具有增强机体免疫力、调节机体生理、预防疾病、促进健康等功能[2],更重要的是产品具有特殊的风味。风味的优劣是人们选择发酵乳制品的基本条件之一,因此发酵乳制品风味的研究对于产品风味的控制和开发新产品具有重要的理论和实践意义。
1 风味形成的机制
食物所产生的气味,一般都由许多种挥发性物质所组成。这些物质形成途径十分复杂,许多反应的机制至今仍不清楚[3]。食品中风味的形成途径大致可以分成生物合成、酶促作用、氧化分解和高温分解作用4种机制(见表1)。
2 发酵乳品风味产生的机制
发酵乳制品的香气成分包括包括从原料乳中移入的物质、加工中各种反应生成的物质以及由微生物制造出来的物质[3]。应用于发酵乳生产的微生物主要有乳酸菌、霉菌、酵母菌等。乳酸菌主要包括乳酸乳球菌(乳酸乳球菌乳亚种、乳酸乳球菌乳脂亚种、乳酸乳球菌双乙酰亚种)、嗜热链球菌、乳杆菌(德氏乳杆菌保加利亚亚种、植物乳杆菌、瑞士乳杆菌等)和明串珠菌属(如肠膜明串珠菌乳脂亚种)等。这些乳酸菌对发酵乳制品风味有决定性的影响,近年来,科研工作人员对酸奶、发酵奶油、干酪等乳制品的风味进行了分析(见表2)。所检测出的有机化合物包括有有机酸、酮、醛、酯、芳香族化合物、含硫化合物等,这些物质是由乳中乳糖、乳脂肪和乳蛋白通过微生物的发酵或酶促反应产生的[5]。
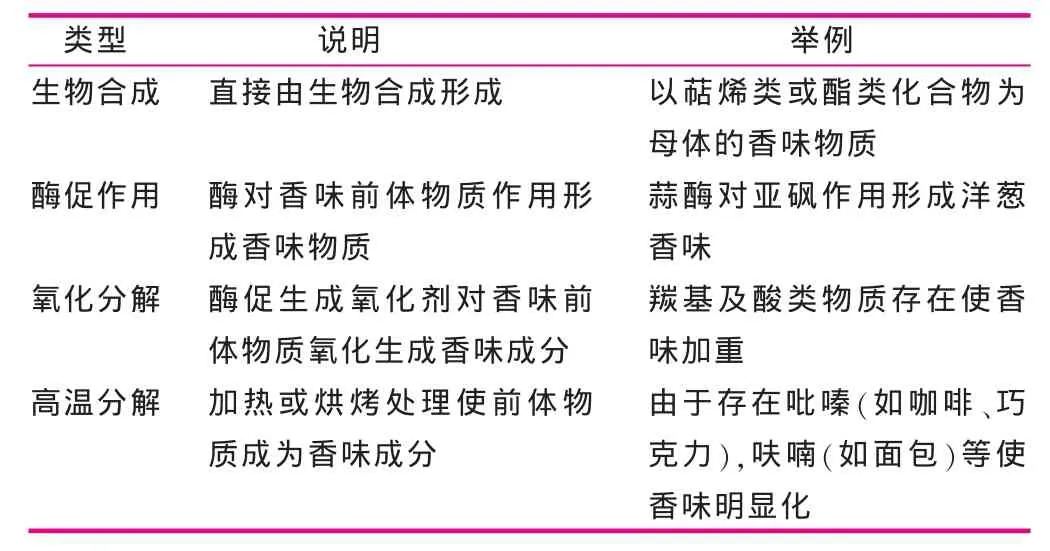
表1 食品香味形成机制的类型[4]

表2 关于发酵乳风味的研究
2.1 乳脂肪源风味物质的产生
乳脂肪是重要的风味物质前体[13],但是有时也能形成异味[14]。乳脂肪水解产生游离脂肪酸,特别是短链、中链脂肪酸,是干酪的主要风味物质。游离脂肪酸还可以代谢产生其他的风味物质,如甲基酮、内酯、酯和仲醇[15]。乳脂肪的代谢途径见图1。
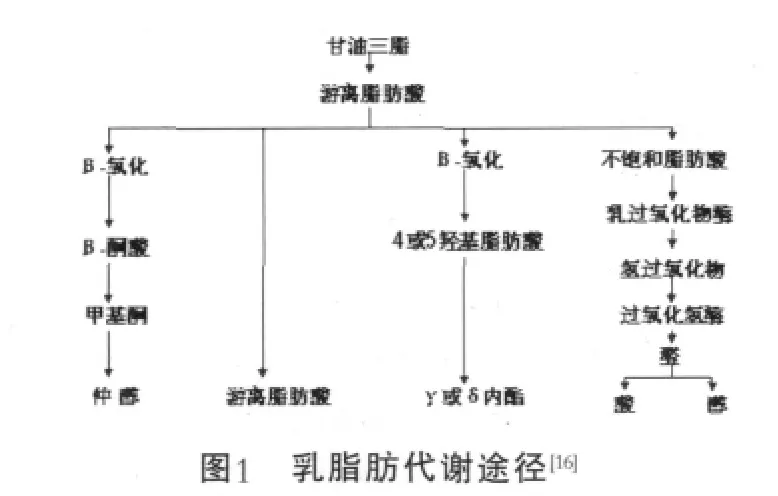
甲基酮是一种重要的风味物质,特别是干酪。甲基酮是由β-氧化途径形成的:脂肪酸经过β-氧化形成β-酮脂酰,经过磷酯酶的作用形成β-酮酸,然后脱羧形成甲基酮[17-18],其中起着分解作用的磷酯酶也叫脱酰酶,acyl-CoA水解酶,acyl-CoA磷酯水解酶和acyl-CoA磷脂酶[19]。磷酯酶存在于过氧化物酶体、线粒体和细胞质,通过调控脂肪酸和acyl-CoAs的浓度来调整脂肪代谢[20]。霉菌的β-氧化体系研究较多,如黑曲霉[21]、娄地青霉菌[22]、锁掷孢酵母[23]、解脂耶氏酵母[24]。细菌的研究较少,如Staphylococcus carnosus[17]、新月柄杆菌[25]等,但是乳酸菌中的β-氧化体系机制还不是很清楚,这将是以后研究的重要内容。
在干酪的风味物质中,除了甲基酮外,还检测到很多内酯,内酯类化合物的前体是羟基脂肪酸和酮酸等,Gouda干酪成熟过程中内酯的形成并不是由于酶的作用,而是一种化学反应,是酯化的羟基脂肪酸经过自身转酯基(羟基攻击酯键)作用释放出内酯[26]。
酯类风味物质的产生主要通过酯化、醇解、酸解和酯交换四种途径[27]:(a)酯化(R1-COOH+R2-OH→R1-COO-R2+H2O);(b)醇解(R1-COO-R2+ R3-OH→R1-COO-R3+R2-OH);(c)酸解(R1-COO-R2+R3-COOH→R3-COO-R2+R1-COOH);(d)酰基转移(酯交换)(R1-COO-R2+R3-COOR4→R1-COO-R4+R3-COO-R2)。乳酸菌通过酯酶催化的酯化和醇解两种酯的合成机制已经在被证实,不过酸解和转酰基还未见报道。山羊和母羊及其干酪中分离的发酵乳杆菌,保加利亚杆菌,鼠李糖乳杆菌,干酪乳杆菌,植物乳杆菌有能力合成短链脂肪酸酯,产物主要是丁酸乙酯和己酸乙酯,合成的机制是酯化[28]。在乙醇存在的水性环境里,Streptococcus thermophilus能够形成乙酸乙酯等短链酯,机制是醇解,乳酸菌酯酶是一种具有转移酶活力的酶,可以选择水(水解)或乙醇(醇解)作为酰基接收器[29],并优先选择甘油一酸酯和甘油二酸酯进行酯的合成。乳酸乳球菌在适宜的条件下,尽管可能存在其他的酯合成途径,不过酯酶起主要作用[30]。乳酸菌酯酶存在着菌种及个体菌株的差异。因此选择合适的发酵剂对于发酵乳制品风味的重要性[31]。
2.2 乳糖和柠檬酸源风味物质的形成
乳酸菌发酵糖产生乳酸、乙醇或者乙酸和二氧化碳等物质,乳酸菌代谢乳糖和柠檬酸的一般途径见图2。
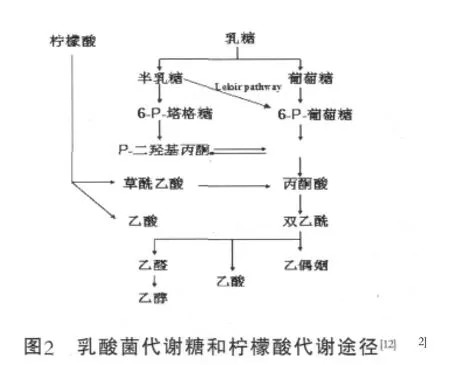
明串株菌属代谢葡萄糖路径首先裂解产生一分子CO2和5P-核酮糖,5P-核酮糖转化为5P-木酮糖之后,由磷酸转酮酶裂解为3-p-甘油醛和乙酰磷酸,这种路径在异型发酵乳杆菌也有[32]。
Lactobacillus helveticus能够同时代谢柠檬酸和乳糖,柠檬酸能够刺激乳糖的消耗和乳酸的产生,代谢方式由同型发酵转变为混合酸模式,主要风味物质是乙酸,其次是琥珀酸,未检测到乙偶姻、双乙酰和丁二酮[33]。Leuconostoc oenos在柠檬酸和葡萄糖同时存在时,柠檬酸刺激了葡萄糖的代谢,细胞内6-p-glucose减少了10倍,同时NADP浓度和ATP的合成量增加[34]。Lactobasillus casei和Lactobacillus plantarum不能把柠檬酸作为能量来源,代谢主要产物是乙酸和乙偶姻,有时也能生成丙酮酸,当柠檬酸耗尽后,转变为乙偶姻[35]。L.lactissubsp.lactis biovar diacetylactis可 以在柠檬酸是唯一碳源的培养条件下生长,代谢产物为丙酮酸、乙酸、乙偶姻和丁二酮[36]。
对于糖的代谢研究,一般常用13cNMP和31PNMR核磁共振光谱的方法,Hugenholtz(2000)对L.lactic糖的代谢和胞外多糖的产生进行了研究,结果表明31PNMR能够察觉到在葡萄糖代谢期间L.lactic瞬间的生理变化[36]。Ramos对Leuconostoc oenos柠檬酸和糖代谢进行了研究,13CNMP可以清晰检测到最终产品的代谢转变,通过研究发现13CNMP和31PNMR核磁共振光谱是研究乳酸菌糖类代谢有效的办法[34]。
2.3 氨基酸源风味物质的产生
乳酸乳球菌代谢氨基酸是通过转氨作用产生α-酮酸,这一步是由转氨酶催化的,主要有芳香族转氨酶、支链转氨酶和天冬氨酸转氨酶[37];α-酮酸进一步由酮酸脱羧酶转化为乙醛,或是由酮酸脱氢酶转化成羧酸,乳酸代谢氨基酸的一般途径见图3。
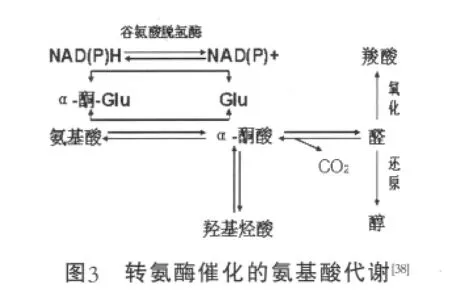
L.Lactis代谢亮氨酸和缬氨酸的产物主要是谷氨酸,说明α-酮戊二酸是氨基的主要接收物质[39],但也有少数乳球菌把丙酮酸或草酰乙酸作为氨基的接收物质[40]。α-酮戊二酸是乳酸菌代谢氨基酸的限制性因素,形成主要有3种途径:一是可以由谷氨酸在谷氨酸脱氢酶作用下脱氢生成;第二途径是柠檬酸-草酰乙酸途径,由柠檬酸和谷氨酸产生,需要柠檬酸盐渗透酵素(CitP),柠檬酸裂解酶(CitL)和天冬氨酸转氨酶(Asp-AT);第三途径是柠檬酸-异柠檬酸途径,利用三羧酸循环的氧化之路,由丙酮酸或柠檬酸氧化产生,需要异柠檬盐脱氢酶和顺乌头酸酶。Tanous通过L. lactissubsp.Diacetylactis获得了柠檬酸盐渗透酵素,柠檬酸裂解酶和天冬氨酸转氨酶,而没有获得异柠檬盐脱氢酶和顺乌头酸酶,证明α-酮戊二酸第二种形成途径是存在的[41]。
Williams对切达干酪中152株乳酸菌在酮戊二酸盐存在的条件下分解蛋白质和氨基酸的能力进行了研究[42]。Liu对71株乳酸菌代谢氨基酸产生的风味物质进行了研究,精氨酸可以被乳酸乳球菌乳亚种代谢,也可被短乳杆菌、发酵乳杆菌等异型发酵杆菌代谢,丝氨酸只能被同型发酵的类干酪乳杆菌类干酪亚种、鼠李乳杆菌和植物乳杆菌代谢[29]。其中短乳杆菌和发酵乳杆菌代谢氨基酸能力强,可以利用丝氨酸、精氨酸、天门冬氨酸、谷氨酸和支链氨基酸,明串珠菌代谢氨基酸能力弱。乳酸菌代谢支链氨基酸能产生类似麦芽味,水果味和汗臭味,芳香族氨基酸产生花、化学试剂和排泄物的味道,天冬氨酸代谢产生类似黄油风味,含硫氨基酸代谢产生类似煮沸的卷心菜、肉和大蒜味道[38]。
3 乳酸菌调控和影响风味形成的途径
3.1 改善乳酸菌的发酵条件
在乳球菌的发酵过程中,芳香物质的生成受到发酵条件的影响,如双乙酰的生成量和稳定性受多方面外界因素的影响,如培养基中的碳、氮源的种类和比例,生长温度,pH,是否存在氧气等[43]。Lactococcus lactisssp.lactis在有氧条件下,双乙酰的生成量远远高于无氧条件下的生成量,氧气的压力达到1×105Pa和2×105Pa时,双乙酰的产生量的最大值分别达到0.39和0.45 mmol/L[44]。氧气的压力达到2×105Pa时,未添加柠檬酸时,双乙酰和乙偶姻最大浓度分别为0.09和0.29 mmol/L,当柠檬酸的添加量由6.2,11.5,26 mmol/L,双乙酰的最大浓度分别为0.45,0.53,0.85 mmol/L,乙偶姻为3.6,4.5,8.5 mmol/L,当柠檬酸的浓度增加时,由柠檬酸转化的双乙酰却在减少[45]。
3.2 基因工程
近年来双乙酰基因调控主要集中在增强丙酮酸向α-乙酰乳酸代谢支路,失活α-乙酰乳酸脱羧酶以便切断α-乙酰乳酸至乙偶姻的代谢支路和抑制双乙酰还原酶的活性,以及这些策略的综合[46]。Henriksen利用丙酮酸甲酸盐裂解酶(PFL),乳酸脱氢酶(LDH)和丙酮酸脱氢酶(PDH)缺陷性菌株对丙酮酸代谢路径进行了调控[47]。Chaves等通过基因敲除技术来构建gtyA基因缺失的突变株,gtyA基因缺陷菌株的苏氨酸醛缩酶活性几乎完全丧失,乙醛的产量达不到检测限[48];而glyA基因过表达菌株的生长速率和产酸能力与其亲本基本相同,乙醛的产量和苏氨酸醛缩酶活性较其亲本却分别增加了86%和65%[49]。
4 展望
发酵乳制品的风味研究是如今的一个热门话题,目前国内对于发酵乳制品的风味研究主要集中在糖和柠檬酸的代谢上,而对于脂肪酸的代谢研究较少,特别是脂肪酸代谢对于发酵乳风味的影响,将是今后的研究重点之一。
[1]骆承庠.乳与乳制品工艺学[M].北京:农业出版社,1992.
[2]李成涛,吕嘉枥.乳酸菌及其发酵乳制品的发展趋势[J].中国酿造, 2005(8):5-7.
[3]丁耐克.食品风味化学[M].北京:中国轻工业出版社,1999:259.
[4]黄梅丽,王俊卿.食品色香味化学[M].北京:中国轻工业出版社,2008:262-263
[5]宋焕禄.食品风味化学[M].北京:化学工业出版社,2008:79-80
[6]王琴,朱小红,任远庆,等.不同乳酸菌及其组合发酵乳的产香特性分析[J].食品工业科技,2008(6):73-76.
[7]王冰.应用复合乳酸菌开发天然酸奶香基[J].香料香精化妆品,2008 (3)):49-52
[8]汪薇,赵文红,白卫东,等.乳酸菌发酵制备天然奶味香精的研究[J].食品与发酵工业,2010(10):191-195.
[9]AWAD S,AHMED N,SODA M E.Evaluation of Isolated Starter Lactic Acid Bacteria in Ras Cheese Ripening and Flavour Development [J].Food Chemistry,2007,104(3):1192-1199.
[10]AYAD E H E,VERHEUL A,BRUINENBERG P,et al.Starter Culture Development for Improving the Flavour of Proosdij-type Cheese[J].International Dairy Journal,2003,13(2-3):159-168.
[11]ASTERI I A,ROBERTSON N,KAGKLI D M,et al.Technological and Flavour Potential of Cultures Isolated from Traditional Greek Cheeses–A pool of Novel Species and Starters[J].International Dairy Journal,2009,19(10):595-604.
[12]MARILLEY L.Flavours of Cheese Products:Metabolic Pathways, Analytical Tools and Identification of Producing Strains[J].International Journal of Food Microbiology,2004,90(2):139-159.
[13]FODA M S,KHAFAGY E Z,EL-DIN S M.Production of L-asparaginase by Pseudomonas ovalis[J].Zentralbl Bakteriol Parasitenkd Infektionskr Hyg,1974,129(6):525-532.
[14]SURIYAPHAN O,DRAKE M A,CHEN X Q,et al.Characteristic Aroma Components of British Farmhouse Cheddar Cheese[J].J Agric Food Chem,2001,49(3):1382-1387.
[15]FOX P F WALLACE J M.Formation of Flavor Compounds in Cheese[J].Adv Appl Microbiol,1997,45:17-85.
[16]MOLIMARD P,SPINNLER H E.Review:Compounds Involved in the Flavor of Surface Mold-ripened Cheeses:Origins and Properties [J].Journal of Dairy Science,1996,79(2):169-184.
[17]ENGELVIN G,FERON G,PERRIN C,et al.Identification of Beta-oxidation and Thioesterase Activities in Staphylococcus carnosus 833 Strain[J].FEMS Microbiol Lett,2000,190(1):115-120.
[18]MCSWEENEY F K,MURPHY E S.Criticisms of the Satiety Hypothesis as an Explanation for Within-session Decreases in Responding[J].J Exp Anal Behav,2000,74(3):347-361.
[19]MARY P,CHIHIB N E,CHARAFEDDINE O,et al.Starvation Survival and Viable but Nonculturable States in Aeromonas hydrophila[J].Microb Ecol,2002,43(2):250-258.
[20]VERONIKA P,HANA P,JIRI P,et al.Combined Exposure of Japanese Quails to Cyanotoxins,Newcastle Virus and Lead:Oxidative Stress Responses[J].Ecotoxicol Environ Saf,2011,74(7):2082-2090.
[21]BALTAZAR M F,DICKINSON F M,RATLEDGE C.Oxidation of Medium-chain acyl-CoA Esters by Extracts of Aspergillus niger:Enzymology and Characterization of Intermediates by HPLC[J].Microbiology,1999,145(Pt 1):271-278.
[22]CHALIER P CROUZET J.Methyl Ketone Production from Copra Oil byPenicillium roquefortiSpores[J].Food Chemistry,1998,63(4):447-451.
[23]GILLES F H,GOMEZ I G.Developmental Neuropathology of the Second Half of Gestation[J].Early Hum Dev,2005,81(3):245-253.
[24]ANNE G,GRUBERG L.Platelet Glycoprotein IIb/IIIa Inhibitors during Percutaneous Coronary Interventions:A Pharmacological and Clinical Review[J].Expert Opin Pharmacother,2004,5(2):335-348.
[25]O'CONNELL M H S,SHAPIRO L.Fatty Acid Degradation inCaulobacter crescentus[J].Journal of Bacteriology,1986,168(1):49-54.
[26]ALEWIJÓN M,SMIT B A,SLIWINSKI E L,et al.The Formation Mechanism of Lactones in Gouda Cheese[J].International Dairy Journal,2007,17(1):59-66.
[27]HOLLAND R,LIU S Q,CROW V L,et al.Esterases of Lactic Acid Bacteria and Cheese Flavour:Milk Fat Hydrolysis,Alcoholysis and Esterification[J].International Dairy Journal,15(6-9):711-718.
[28]ABEIJ N MUKDSI M C,MEDINA R B,ALVAREZ M D F,et al. Ester Synthesis by Lactic Acid Bacteria Isolated from Goat’s and Ewe’s Milk and Cheeses[J].Food Chemistry,2009,117(2):241-247.
[29]LIU S Q,HOLLAND R,CROW V L.The Potential of Dairy Lactic Acid Bacteria to Metabolise Amino Acids via Non-transaminating Reactions and Endogenous Transamination[J].International Journal of Food Microbiology,2003,86(3):257-269.
[30]NARDI M,FIEZ-VANDAL C,TAILLIEZ P,et al.The EstA Esterase is Responsible for the Main Capacity ofLactococcus lactisto Synthesize Short Chain Fatty Acid Esters invitro[J].J Appl Microbiol, 2002,93(6):994-1002.
[31]OLISZEWSKI R,MEDINA R B,GONZALEZ S N,et al.Esterase Activities of Indigenous Lactic Acid Bacteria from Argentinean Goats’Milk and Cheeses[J].Food Chemistry,2007,101(4):1446-1450.
[32]COGAN T M,JORDAN K N.Metabolism of Leuconostoc Bacteria [J].Journal of Dairy Science,1994,77(9):2704-2717.
[33]TORINO M I,TARANTO M P,FONT DE VALDEZ G.Citrate Catabolism and Production of Acetate and Succinate byLactobacillus helveticusATCC 15807[J].Applied Microbiology and Biotechnology, 2005,69(1):79-85.
[34]RAMOS A,SANTOS H.Citrate and Sugar Cofermentation in Leuconostoc oenos,a(sup13)C Nuclear Magnetic Resonance Study[J].Appl Environ Microbiol,1996,62(7):2577-2585.
[35]MACCIOLA V,CANDELA G,DE LEONARDIS A.Rapid Gaschromatographic Method for the Determination of Diacetyl in Milk, Fermented Milk and Butter[J].Food Control,2008,19(9):873-878.
[36]HUGENHOLTZ J,LOOIJESTEIJN E,STARRENBURG M,et al. Analysis of Sugar Metabolism in an EPS ProducingLactococcus lactisby 31P NMR[J].Journal of Biotechnology,2000,77(1):17-23.
[37]YVON A M,GROSS D J,WADSWORTH P.Antagonistic Forces Generated by Myosin II and Cytoplasmic Dynein Regulate Microtubule Turnover,Movement,and Organization in Interphase Cells[J]. Proc Natl Acad Sci U S A,2001,98(15):8656-8661.
[38]ARDO Y.Flavour Formation by Amino Acid Catabolism[J]. Biotechnology Advances,2006,24(2):238-242.
[39]ZIADIM,BERGOTG,COURTINP,etal.AminoAcid Catabolism byLactococcus lactisduring Milk Fermentation[J].International Dairy Journal,2010,20(1):25-31.
[40]ENGELS W J M,ALTING A C,ARNTZ M M T G,et al.Partial Purification and Characterization of Two Aminotransferases fromLactococcus lactissubsp.cremorisB78 Involved in the Catabolism of Methionine and Branched-chain Amino Acids[J].International Dairy Journal,2000,10(7):443-452.
[41]TANOUS C,GORI A,RIJNEN L,et al.Pathways for-ketoglutarate Formation by Lactococcus lactis and Their Role in Amino Acid Catabolism[J].International Dairy Journal,2005,15(6-9):759-770.
[42]WILLIAMS A G,NOBLE J BANKS J M.Catabolism of Amino Acids by Lactic Acid Bacteria Isolated from Cheddar Cheese[J].International Dairy Journal,2001,11(4-7):203-215.
[43]杨丽杰,王俊沪.乳酸乳球菌中双乙酰代谢支路的调控[J].中国乳品工业,2004(5):24-29.
[44]BOUMERDASSI H,MONNET C,DESMAZEAUD M,et al.Effect of Citrate on Production of Diacetyl and Acetoin byLactococcus lactis ssp.lactisCNRZ 483 Cultivated in the Presence of Oxygen[J].Journal of Dairy Science,1997,80(4):634-639.
[45]BOUMERDASSI H,MONNET C,DESMAZEAUD M,et al.Isolation and Properties ofLactococcus lactissubsp.lactisBiovar Diacetylactis CNRZ 483 Mutants Producing Diacetyl and Acetoin from Glucose [J].Appl Environ Microbiol,1997,63(6):2293-2299.
[46]郑应福,阚振荣,赵春海.高产双乙酰乳球菌的研究进展[J].中国生物工程杂志,2005(1):186-189.
[47]HENRIKSENCM,NILSSOND.RedirectionofPyruvate Catabolism inLactococcus lactisbySelection of Mutantswith Additional Growth Requirements[J].Applied Microbiology and Biotechnology, 2001,56(5-6):767-775.
[48]CHAVES A,FERNANDEZ M,LERAYER A,et al.Metabolic Engineering of Acetaldehyde Production byStreptococcus thermophilus[J]. Applied and Environmental Microbiology,2002,68(11):5656-5662.
[49]韩希妍,孙大庆,相丽,等.乳酸菌风味代谢物质的基因调控[J].微生物学报,2007(6):1105-1109.
Effect of Lactic acid bacteria on flavor of fermented dairy products
LI Liang1,2,MA Ying1
(1.School of Food Science and Engineering,Harbin Institute of Technology,Harbin 150090,China;2.Food College, Northeast Agricultural University,Harbin 150030,China)
Lactic acid bacteriahave a decisive influence on the flavor of fermented dairy products,Lactic acid bacteriametabolism of fats,carbohydrates and proteins produced flavor substances.This article summarized mechanism research thatLactic acid bacteriametabolism of fat,sugar,citric acid and amino acid to produce aromatic substances
Lactic acid bacteria;flavor;mechanism
Q939.11+7
B
1001-2230(2012)05-0042-04
2011-12-26
李良(1981-),男,讲师,研究方向为食品科学。
马莺
