miR-567通过调控CDK8在NSCLC增殖、迁移和细胞周期中的作用及其临床相关性研究*
2024-02-21李海洋赵振山郑爱民郝孟辉田发明
李海洋,赵振山,李 静,戎 瑶,郑爱民,郝孟辉,田发明
1.开滦总医院肿瘤内科,河北唐山 063000;2.开滦总医院胸外科,河北唐山 063000;3.唐山市人民医院肿瘤科,河北唐山 063000;4.华北理工大学基础医学院,河北唐山 063000
肺癌是威胁人类生命的主要癌症类型,非小细胞肺癌(NSCLC)占所有肺癌病例的85%,且NSCLC患者诊断时通常为处于疾病晚期[1]。与小细胞肺癌相比,NSCLC对化疗或放疗相对不敏感,虽然有关NSCLC治疗的抗癌药物的研究很多,但因耐药、转移、复发等原因导致NSCLC患者预后差,存活率低[2]。因此,迫切需要发现新的生物标志物和检测靶点用于NSCLC患者的诊断和治疗。据研究报道,NSCLC中存在微小RNA(miRNA)失调,miRNA 可以通过靶向致癌基因或抑癌基因,调节细胞增殖、细胞周期和细胞凋亡来参与癌症的发展[3-5]。miR-567由位于染色体3q13.2上的miR-567基因编码[6]。最近研究表明,miR-567在乳腺癌和骨肉瘤等癌症中发挥重要作用[7-8]。然而,miR-567在 NSCLC中的作用仍未阐明。先前的研究结果证实了miR-567可通过调节多种细胞周期蛋白,细胞周期蛋白依赖激酶(CDKs)影响细胞增殖和细胞周期[9]。细胞周期蛋白依赖性激酶8(CDK8)是CDKs的重要成员,参与调节多种癌症信号通路的致癌基因[10]。但 CDK8 在 NSCLC 中的表达模式和功能至今仍未明确,miR-567是否介导CDK8参与NSCLC进程尚未可知。本研究旨在探讨miR-567对A549 NSCLC细胞增殖、迁移和凋亡的潜在调控作用,并明确miR-567对CDK8基因的靶向调节作用,阐明可能有助于NSCLC治疗的相关分子机制,以期为临床NSCLC的诊断和药物治疗提供新的靶位点和分子治疗方向。现报道如下。
1 资料与方法
1.1一般资料 收集2020年8月至2022年8月于开滦总医院进行手术的40例NSCLC患者的肿瘤组织及其癌旁组织(距肿瘤边缘≥3 mm),患者此前均未接受放疗和化疗。取样后立即放在冻存管并置于液氮中。40例NSCLC患者中男30例,年龄为(57.46±7.14)岁;女10例,年龄为(59.23±5.86)岁。所有患者术前均已签署知情同意书。本研究在开滦总医院伦理委员会的监督下获得批准和实施。
1.2仪器与试剂 人NSCLC细胞株A549和H1975均购自American type culture collection(ATCC); RevertAidTM第一链 cDNA Synthesis试剂盒(货号:K1621)购自赛默飞世尔科技(中国)有限公司;SYBR®Green Pro Taq HS 预混型qPCR试剂盒(货号:AG11701)购自湖南艾科瑞生物工程有限公司;PCR引物由上海生工生物工程有限公司合成;DMEM(货号:PM150210B)和0.25%胰酶消化液(货号:PB180223)购自武汉普诺赛生命科技有限公司; miR-NC mimic、miR-567 mimic、oe-NC和oe-CDK8质粒由广州锐博生物科技有限公司合成;荧光素酶报告质粒由上海吉玛基因公司提供;荧光素酶报告基因检测试剂盒(货号:RG027)购自上海碧云天生物技术有限公司;Trizol 试剂(货号:15596-026)、Lipofectamine 2000(货号:11668-027)购自美国Invitrogen;CCK-8(货号:ck04)购自日本同仁化学研究所;Transwell 小室(规格:3422)购自美国Corning公司;细胞周期检测试剂盒(货号:E-CK-A351)购自武汉伊莱瑞特生物科技有限公司。高速离心机(型号:Microfuge16)购自贝克曼库尔特国际贸易(上海)有限公司;实时荧光定量PCR仪(型号:CFX Connect)购自美国BIO-RAD;显微镜(型号:BX41)购自日本Olympus;Spark多功能酶标仪购自瑞士TECAN;流式细胞仪(型号:DxFLEX)购自贝克曼库尔特国际贸易(上海)有限公司。
1.3方法
1.3.1实时荧光定量PCR(qRT-PCR)检测基因表达水平 使用 Trizol 试剂提取肿瘤组织或培养细胞中的总RNA,并使用酶标仪检测所有样品的RNA浓度和纯度。提取的RNA样品应保存在-80 ℃。使用 RevertAidTM第一链 cDNA 合成试剂盒合成cDNA。然后根据SYBR®Green Pro Taq HS 预混型 qRT-PCR试剂盒的说明书,按照说明书配制 qRT-PCR 反应体系。内参基因甘油醛-3-磷酸脱氢酶(GAPDH)应用于计算 mRNA 的最终表达量,计算方法为 2-ΔΔCt。引物序列为miR-567上游:5′-ACAGGACACTATGTTCTTCCAGG-3′;下游:5′-AAAGT TTTGCATGTTCTTCCAGT-3′。CDK8上游:5′-GCC GGTTGTCAAATCCCTTAC-3′;下游:5′-TGTGAC TGCTGTCTTGATTCCCT-3′。GAPDH上游:5′-T TGCAACCGGGAAGGAAATG-3′;下游:5′-TGGA ATTTGCCATGGGTGGA-3′。
1.3.2细胞培养 从4 ℃取出10%胎牛血清的DMEM培养基,迅速取出冻存的A549和H1975细胞,加入含5 mL DMEM培养基的离心管中离心,弃掉上清,加入含10%血清的新鲜培养基,置于37 ℃、5%CO2的细胞培养箱中培养。当细胞生长至70%~80%时,使用0.25%胰酶消化液消化细胞,加入含10%血清的培养基终止消化,离心沉淀细胞,去除胰酶,加入新鲜培养基重悬细胞沉淀。根据培养基颜色变化更换培养基。细胞的传代培养应在对数生长期进行。
1.3.3细胞转染 设计合成 miR-567 的反义寡核苷酸序列(miR-567 mimic)和过表达 CDK8 的质粒(oe-CDK8)。在 6 孔板中培养 A549 和 H1975 细胞,待细胞生长至 50%~60% 融合度时,将细胞分成4组:miR-NC mimic 组(转染 miR-NC mimic)、miR-567 mimic 组(转染 miR-567 mimic)、miR-567 mimic+oe-NC 组(转染 miR-567 mimic 和 oe-NC)和 miR-567 mimic+oe-CDK8 组(转染 miR-567 mimic 和 oe-CDK8)。采用 Lipofectamine 2000 瞬时转染技术,按照说明书的步骤进行转染。
1.3.4CCK-8法检测细胞增殖水平 将来自不同处理组的A549和H1975细胞接种到96孔板中,每孔细胞数目为2×103个,并在含有5%CO2的37 ℃培养箱中培养。在第0、24、48或72小时将10 μL CCK-8溶液小心添加到细胞中(注意不要在孔中生成气泡)。在培养箱中继续孵育2 h后,使用酶标仪在450 nm 处测定吸光度(A)值。
1.3.5Transwell法检测细胞迁移能力 将带有8 μm孔径聚碳酸酯膜过滤器的Transwell小室置于标准24孔细胞培养板中。然后,在上室加入200 μL(含5×105个细胞)细胞悬液(无血清DMEM培养基重悬)。在下室加入800 μL含10%胎牛血清的DMEM。将24孔板在含有5%CO2的37 ℃培养箱中培养24 h。培养结束后,下室细胞用4%多聚甲醛固定并用结晶紫染色。PBS清洗3次后在显微镜下拍照并计数。
1.3.6流式细胞术检测细胞周期 按照细胞周期检测试剂盒的说明书操作。将不同处理后的对数生长期 A549 和 H1975 细胞进行胰酶消化成单细胞悬液,用预冷的PBS溶液漂洗2次,以1 500 r/min 的速度离心 10 min,弃去上清后,加入 100 μL 稀释好的 binding buffer 重悬细胞,然后分别加入 5 μL RNAase PI 染色液。在避光条件下,在 37 ℃ 孵育 30 min后,加入 500 μL 稀释好的 binding buffer。将样品使用单细胞筛网过滤后,使用流式分析仪进行检测。
1.3.7生物信息学预测 为了预测 miR-567可能的靶基因,使用了3个在线软件程序,包括 TargetScan(http://www.TargetScan.org/),miRDB(http://miRDB.org/miRDB/)和 Diana Tools(http://Diana.imis.athena-innovation.gr/dianatools/)进行生物信息学预测。
1.3.8荧光素酶报告基因实验 构建包含miR-567与CDK8 3′UTR区结合位点的正常型CDK8载体质粒和突变结合位点的 CDK8-MUT 载体质粒。将处于对数生长期的A549和H1975细胞以3×105/孔的密度接种于24孔板中,分别将正常型CDK8或突变型CDK8同miR-567 mimics 或 miR-567 NC 一起转染至A549和H1975细胞。48 h后,将细胞裂解并检测荧光素酶活性。计算公式:相对荧光素酶活性=萤火虫荧光素酶活性/海肾荧光素酶活性。
1.4统计学处理 使用GraphPad Prism 8.0软件统一处理数据。两个参数之间的相关性分析采用Pearson相关系数法,计算相关系数(r)和双尾P值。多组间的比较采用单因素方差分析(One-way ANOVA)和 Tukey 多重比较,两组之间的比较采用t检验进行。P<0.05表示差异有统计学意义。
2 结 果
2.1NSCLC患者肿瘤组织中miR-567和CDK8表达变化 qRT-PCR结果显示,与癌旁组织比较,肿瘤组织中miR-567表达降低,CDK8表达升高,差异有统计学意义(P<0.05),见图1A。Pearson相关系数分析结果显示,在肿瘤组织中miR-567和CDK8表达呈负相关(P<0.05)。见图1B。

注:A为qRT-PCR检测miR-567和CDK8表达;B为Pearson相关系数分析miR-567和CDK8表达相关性;与癌旁组织比较,*P<0.05。
2.2过表达miR-567抑制NSCLC细胞CDK8表达 qRT-PCR结果显示,在A549和H1975细胞中与miR-NC mimic组比较,miR-567 mimic组miR-567表达升高,CDK8表达降低,差异有统计学意义(P<0.05)。见图2。

注:A为qRT-PCR检测miR-567表达;B为qRT-PCR检测CDK8表达;与miR-NC mimic组比较,*P<0.05。
2.3过表达miR-567抑制NSCLC细胞增殖和细胞迁移 CCK-8法结果显示,在A549和H1975细胞中与miR-NC mimic组比较,miR-567 mimic组细胞增殖水平降低,差异有统计学意义(P<0.05)。见图3A。Transwell法结果显示,与miR-NC mimic组比较,miR-567 mimic组细胞迁移水平降低,差异有统计学意义(P<0.05)。见图3B。

注:A为CCK-8法检测细胞增殖水平;B为Transwell法检测细胞迁移水平;与miR-NC mimic组比较,*P<0.05。
2.4过表达miR-567阻滞NSCLC细胞周期 流式细胞术结果显示,在A549和H1975细胞中与miR-NC mimic组比较,miR-567 mimic组细胞G1期比例升高,S期比例降低,差异有统计学意义(P<0.05)。见图4。
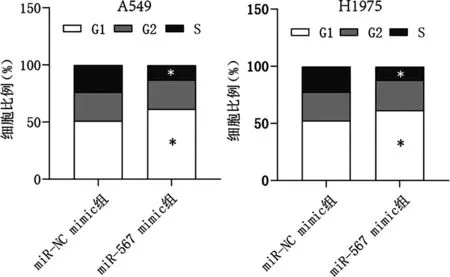
注:与miR-NC mimic组比较,*P<0.05。
2.5miR-567靶向结合CDK8 生物信息学预测结果显示,miR-567与CDK8具有7个连续的碱基结合位点,具有可能的靶向性。见图5A。通过荧光素酶报告基因实验,发现在含有正常型CDK8的A549和H1975细胞中,miR-567 mimic组荧光强度显著低于miR-NC mimic组,差异有统计学意义(P<0.05)。而在突变型CDK8中,miR-567 mimic组和miR-NC mimic组荧光强度比较差异无统计学意义(P>0.05)。见图5B。

注:A为miR-567与CDK8结合位点;B为荧光素酶报告基因实验检测miR-567和CDK8结合情况;与miR-NC mimic组比较,*P<0.05。
2.6过表达CDK8促进NSCLC细胞增殖和细胞迁移 qRT-PCR结果显示,在A549和H1975细胞中,miR-567 mimic+oe-CDK8组miR-567表达与miR-567 mimic+oe-NC组比较,差异无统计学意义(P>0.05),见图6A;在A549和H1975细胞中与miR-567 mimic+oe-NC组比较,miR-567 mimic+oe-CDK8组CDK8表达升高,差异有统计学意义(P<0.05),见图6B。CCK-8法结果显示,与miR-567 mimic+oe-NC组比较,miR-567 mimic+oe-CDK8组细胞增殖水平升高,差异有统计学意义(P<0.05)。见图7A。Transwell法结果显示,与miR-567 mimic+oe-NC组比较,miR-567 mimic+oe-CDK8组细胞迁移水平升高,差异有统计学意义(P<0.05)。见图7B。
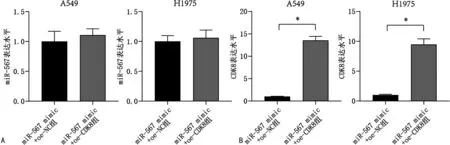
注:A为qRT-PCR检测miR-567表达;B为qRT-PCR检测CDK8表达;与miR-567 mimic+oe-NC组比较,*P<0.05。

注:A为CCK-8法检测细胞增殖水平;B为Transwell法检测细胞迁移水平;与miR-567 mimic+oe-NC组比较,*P<0.05。
2.7过表达CDK8促进NSCLC细胞周期 流式细胞术结果显示,在A549和H1975细胞中与miR-567 mimic+oe-NC组比较,miR-567 mimic+oe-CDK8组细胞G1期比例降低,S期比例升高,差异有统计学意义(P<0.05)。见图8。

注:与miR-567 mimic+oe-NC组比较,*P<0.05。
3 讨 论
目前,NSCLC患者的预后情况并不乐观,转移和复发是NSCLC患者死亡的主要原因,而临床治疗效果的提高在很大程度上依赖于新靶点和生物标志物的发现[11]。不同类型的肺癌治疗方法不同,根据相关生物标记物确定亚型,进而实现精准靶向治疗。
miRNA是基因转录后水平上重要的表达调控因子[12],大约30%的mRNA由miRNAs进行调控。多项研究结果显示,miRNAs在肿瘤细胞生长、增殖和分化等多种生物和病理过程中发挥了重要作用,miRNAs可作为癌症等多种疾病的生物标志物和潜在治疗靶点[13]。并且有研究表明,miRNAs的异常表达通过靶向抑癌基因或癌基因参与了癌症的进展[14]。因此,miRNAs可作为致癌基因或肿瘤抑制因子,并有望成为临床诊断的候选生物标志物,包括识别癌症类型或肿瘤亚型。有研究表明,miRNA 的异常表达通过靶向肿瘤抑制基因或癌基因参与癌症进展[15]。有研究表明,miR-567可以抑制骨肉瘤细胞的增殖、迁移和侵袭,从而缓解骨肉瘤的疾病发展[8]。关于miR-567在NSCLC中作为致癌基因或抑癌基因的生物学作用的研究鲜见报道。基于综合生物信息学分析,有一项研究预测miR-567可能是NSCLC的推定生物标志物[16]。因此,本研究探讨了miR-567在NSCLC细胞中的作用,并阐明了其在NSCLC治疗中可能相关的分子机制。
本研究检测miR-567在NSCLC肿瘤组织中的表达模式,初步探讨miR-567参与NSCLC发生发展的可能性。本研究结果表明,与癌旁组织相比,miR-567表达在NSCLC肿瘤组织中显著降低。异常的细胞增殖和细胞周期状态是癌症的诱发因素之一[17]。本研究分析发现,过表达miR-567显著抑制NSCLC细胞增殖和迁移,并阻滞细胞周期。这一结果表明,miR-567可能是NSCLC细胞的肿瘤抑制因子,并可能成为NSCLC的新型的生物标志物。一项研究发现,miR-567通过阻止胃癌细胞的增殖发挥肿瘤抑制因子的作用[9]。LIU等[8]发现miR-567上调对骨肉瘤细胞增殖具有抑制作用,表明其作为肿瘤抑制因子的功能。细胞周期由不同的阶段组成,包括静息Gap1阶段(G1期)、DNA合成阶段(S期)、Gap2阶段(G2期)和有丝分裂阶段(M期)[18]。本研究结果表明,miR-567上调显著增加G1期的细胞群,显著减少S期细胞群。表明miR-567通过诱导细胞周期停滞抑制NSCLC细胞的增殖和迁移。
CDKs 是参与细胞周期和细胞增殖控制的核心蛋白,CDKs失调会破坏正常的细胞分化、增殖、细胞周期和细胞凋亡,从而导致疾病和癌症的发生[19]。CDK8被认为是一种抑制细胞凋亡的致癌基因,异常的CDK8表达通常与癌症发展有关[20]。近期研究表明,CDK8可通过β-catenin信号轴促进胰腺癌细胞的血管生成,从而加速肿瘤发展[21]。本研究发现,CDK8在NSCLC患者的肿瘤组织中表达上调,miR-567与CDK8表达呈负相关。miRDB 程序及TargetScan 和 Diana Tools 等生物信息学预测发现CDK8是miR-567的靶基因,说明miR-567可能对于CDK8具有靶向负调控作用。为了证实这一推测,本研究评估了NSCLC 细胞中miR-567过表达后CDK8的表达水平,以及过表达CDK8 对于miR-567表达的影响。结果显示,miR-567 的过表达显著降低了 CDK8 mRNA 的表达水平,同时,CDK8过表达逆转了上调miR-567对NSCLC 细胞增殖、迁移的抑制作用及对细胞周期的阻止作用。这些发现表明CDK8是miR-567 的靶基因,miR-567通过靶向负调控CDK8的表达抑制NSCLC的发生发展。
综上所述,本研究拟通过细胞转染技术使人NSCLC细胞株A549和H1975过表达miR-567,研究miR-567对A549和H1975细胞增殖、迁移和细胞周期的作用,并进一步验证miR-567与CDK8基因的靶向调节关系,探讨miR-567的作用机制,从而为NSCLC临床治疗提供新的靶点,可为相关后续研究提供一定的实验依据和技术支持。
