铜绿假单胞菌单PilZ结构域蛋白PA0012和PA4324突变体生物学表型的初步筛选
2024-01-26赵朔,盛硕
赵 朔,盛 硕
(1.遵义医科大学 基础医学院电镜室,贵州 遵义 563099;2.华南农业大学 群体微生物研究中心/广东省微生物信号与作物病害防控重点实验室,广东 广州 510642;3.遵义医科大学 基础药理教育部重点实验室暨特色民族药教育部国际合作联合实验室, 贵州 遵义 563099)
铜绿假单胞菌是一种常见的革兰阴性致病菌,其对寄主的感染可分为急性感染和慢性感染[1],从这两种不同的感染中分离得到的铜绿假单胞菌具有不同的毒性特征[2]。环二鸟苷酸(cyclic diguanosine monophosphate,c-di-GMP)是1987年在木葡糖酸醋杆菌中被首次鉴定的一种能够激发纤维素合成酶构象改变的小分子[3]。随后的研究发现c-di-GMP不仅仅存在于这一种细菌中,也广泛存在于其它细菌中,并且对细菌生物被膜的形成有重要调控作用[4-5]。
C-di-GMP能够由含有GGDEF结构域的二鸟苷酸环化酶(diguanylate cyclase, DGC)催化两分子三磷酸鸟苷(guanosine triphosphate, GTP)环化形成,以及被含有EAL或HD-GYP 结构域的磷酸二酯酶(phosphodiesterase, PDE)特异性地水解。在细菌体内,DGC和PDE紧密地控制着c-di-GMP的浓度,c-di-GMP的受体能通过结合c-di-GMP并与其它蛋白相互作用而导致细胞内的生理变化。含PilZ结构域的蛋白是最早被发现的c-di-GMP受体,并且与其它受体蛋白相比较,PilZ结构域蛋白与c-di-GMP结合能力最强,是最广泛存在于细菌中的c-di-GMP受体。PilZ结构域中的2个基序对c-di-GMP的结合有重要作用,(D/N)xSxxG基序能够结合c-di-GMP,RxxxR基序能够在c-di-GMP附近形成环状,且2个精氨酸(R)能够与c-di-GMP形成氢键。
单PilZ结构域蛋白是所有PilZ结构域蛋白中数量最多的一类,它们通常由80~160个氨基酸组成。单PilZ结构域蛋白也被认为是最难研究的一类PilZ结构域蛋白,相比较于多结构域PilZ蛋白,它们缺少其它结构域为鉴定蛋白功能提供线索。铜绿假单胞菌PAO1的基因组一共编码有8个含PilZ结构域的蛋白,其中PilZ (PA2960),HapZ (PA2799),MapZ (PA4608),PA0012和PA4324是单PilZ结构域蛋白,PilZ(PA2960)是铜绿假单胞菌PAO1产生四型菌毛(T4P)所必要的蛋白[6]。本课题组前期研究发现HapZ(PA2799)能够直接与组氨酸激酶SagS相互作用,阻断SagS向其下游的HptB的磷酸转移[7],MapZ(PA4608)能够跟甲基转移酶CheR1结合,并抑制趋化受体蛋白PctA的甲基化,通过改变趋化受体的甲基化程度来调节细菌鞭毛转动方向变化的频率[8]。 然而,PA4324和PA0012这2个单PilZ结构域蛋白的功能仍然不清楚,这也是本论文的研究主题。
C-di-GMP的典型作用是作为“黏合剂”将2个结构域或者蛋白的距离拉得更近,造成构象的改变,从而触发下游的信号传导。不同的PilZ结构域蛋白与c-di-GMP的结合方式也不一样[9],比如MapZ(PA40608)蛋白与插入式的c-di-GMP结合,而PlzD(VCA0042)蛋白与c-di-GMP单体结合。此外,PilZ结构域还可以通过构象改变灵活地转换与c-di-GMP的结合方式。然而,也有不能与c-di-GMP结合的含PilZ结构域的蛋白,比如铜绿假单胞菌中的蛋白PA2960[10]。因此,不同的PilZ结构域蛋白突变可能触发不同的c-di-GMP信号传导方式,进而影响不同的生物学表型。本论文主要内容为研究PA4324和PA0012对铜绿假单胞菌生物学表型的影响,本研究首先利用同源重组的方法进行基因敲除,得到突变体菌株ΔPA0012和ΔPA4324,随后,以野生型菌株PAO1为对照,对2个突变体进行表型筛选,本研究发现这2个基因的突变导致铜绿假单胞菌的表型变化非常相似,然而,在转录水平上2个蛋白突变导致的差异基因却有明显差异,这些基因所涉及的信号通路也不尽相同。本项研究将能够使研究者更进一步理解铜绿假单胞菌中单PilZ结构域蛋白功能的多样性和重要性,为进一步研究它们调控致病性的机理提供基础。
1 材料与方法
1.1 材料
1.1.1 菌株、质粒和培养条件 本文用到的所有菌株和质粒均列在表1中。编码完整或部分蛋白质的DNA片段都是根据已知的铜绿假单胞菌PAO1基因组序列设计引物,利用PCR手段以该菌的基因组DNA为模板扩增得来的,本文中PCR和分子克隆所使用的所有引物均在表2中进行了总结。质粒都是由标准的分子克隆技术构建完成的,除非有特殊说明。基因敲除使用同源重组的方法。

表1 菌株和质粒汇总

表2 PCR和分子克隆所用引物
1.1.2 主要试剂 实验中用到的抗生素如卡那霉素(Kan) 、羧苄青霉素(Cb)、庆大霉素(Gen)等均购自生工生物工程(上海)股份有限公司;LB等培养基和琼脂粉用的是BD品牌产品;T4 连接酶、PCR 反应高保真酶以及各种限制性内切酶用的是NEB品牌产品;质粒提取试剂盒、琼脂糖凝胶DNA 回收试剂盒用的是Axygen品牌产品;PCR 反应Taq酶购自全式金生物技术有限公司;实验所用到的引物合成和DNA片段的一代测序均由北京擎科新业生物技术有限公司提供。
1.2 方法
1.2.1 基因敲除突变体的构建 构建基因敲除突变菌株用的是同源重组的实验方法,选用的是自杀质粒pK18GT。先将基因上下游同源臂融合并克隆到pK18GT上,转化入感受态细胞DH5α中,随后将DH5α与PAO1和含有辅助质粒的大肠杆菌进行三亲杂交,筛选出被pK18GT重组质粒插入到基因组上的铜绿假单胞菌,再在蔗糖-LB平板上划线,挑取缺失PA0012片段大小的PCR产物所对应的单菌落。
1.2.2 生长曲线实验 挑取单菌落接种至LB液体培养基中,37 ℃,200 r/min过夜培养,随后将所得菌液稀释至OD600≈0.2。取稀释好的100 μL菌液放进全自动生长曲线仪设置37 ℃,150 r/min,测生长曲线48 h。
1.2.3 群集运动实验 该实验使用的培养基是营养肉汤培养基。挑取单菌落接种至LB液体培养基中,37 ℃,200 r/min过夜培养。随后将摇好的种子液接种至现配好的营养肉汤固体培养基平板中央,37 ℃正置培养过夜。
1.2.4 线虫慢杀死实验 该实验使用的培养基是NG固体培养基。挑约30头L4期的雌雄同体线虫到涂有铜绿假单胞菌的NG平板上,20~25 ℃正置培养。在开始培养之后大约12~24 h开始计数,一共观察7 d,每天取2~3个时间点计数剩余的线虫。
1.2.5 脱脂牛奶琼脂平板实验 该实验使用的培养基是脱脂奶粉固体培养基。挑取单菌落接种至LB液体培养基中,37 ℃,200 r/min过夜培养。随后,吸取20 μL菌液加入提前打好孔的脱脂奶粉固体平板的孔中,37 ℃过夜培养,然后测量每个孔周围的蛋白降解圈直径。
1.2.6 生菜感染实验 该实验使用的培养基是LB液体培养基,重悬菌体使用的是10 mmol/L硫酸镁溶液。挑取单菌落接种至LB液体培养基中,37 ℃,200 r/min过夜培养。随后用硫酸镁溶液重悬,取10 μL菌液注射进洁净的生菜叶片中,同时注射硫酸镁溶液作为对照,将接种过的生菜叶片放在无菌的盒子中放入30 ℃培养5 d,每天观察1次。
1.2.7 RNA测序分析 首先,对总RNA的样品检测, RNA质量检测达标以后,进行文库的构建,主要包括去除rRNA,打断mRNA并合成双链cDNA,给纯化后的cDNA加上测序接头并进行片段大小选择,然后将选择好的片段进行PCR扩增,并将所得PCR产物纯化,即得了所需文库。最后,对文库进行质量检测,将库检合格的文库进行HiSeq/MiSeq测序。将所得到的原始数据参照铜绿假单胞菌的参考基因组进行生物信息学的分析。
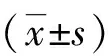
2 结果
2.1 PA0012和PA4324氨基酸序列比对 本研究将PA0012、PA4324、MapZ和HapZ这4个铜绿假单胞菌中单PilZ结构域蛋白的氨基酸序列进行比对,其中MapZ和HapZ是c-di-GMP结合位点已知的2个蛋白。比对结果发现它们具有高度保守的能够与c-di-GMP结合的(D/N)xSxxG基序和RxxxR基序,但其他部分氨基酸序列则存在明显的差异(图1)。

A:4个单PilZ结构域蛋白氨基酸序列比对结果,方框内为最保守的氨基酸位点;B:4个单PilZ结构域蛋白的N端部分氨基酸序列标识,红色星号下的氨基酸为预测的c-di-GMP结合位点。图1 序列比对和序列标识
2.2PA0012和PA4324基因突变不显著影响生长曲线 在进行突变体表型筛选之前,本研究首先进行了生长曲线的测定,以确定这2个基因突变不会影响细菌的生长,本实验采用的是LB液体培养基。用Bioscreen C全自动生长曲线仪所测得的实验结果显示,相比较于野生型菌株PAO1,突变体ΔPA0012和ΔPA4324的生长曲线没有显著性的变化,不接种菌液的LB培养基作为该实验的阴性对照(图2)。该结果表明,铜绿假单胞菌PA0012和PA4324基因突变不会显著性影响细菌的生长,可进行下一步的表型筛选实验。
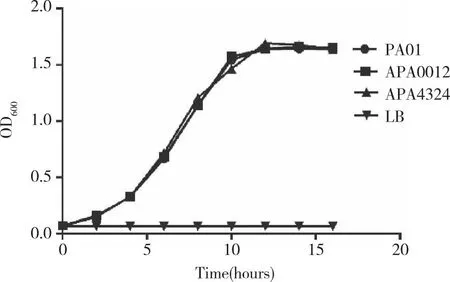
PAO1:野生型;ΔPA0012、ΔPA4324:突变型;LB:培养基。图2 菌株的生长曲线比较
2.3ΔPA0012和ΔPA4324的群集运动能力显著增强 本研究对2个突变体群集运动的能力进行了检测,实验在半固体培养基上进行。实验结果显示,相比较于野生型菌株PAO1,2个突变菌株的群集运动能力均有显著增强,并且均能够被互补(P<0.05,图3A)。表型互补是将蛋白用pUCP18载体在对应的突变菌株中表达,由于互补菌株中含有载体,故在野生型菌株和突变菌株中都加入pUCP18空载体,排除载体影响表型的可能性,在观察群集运动现象的同时,本研究也用尺子对运动区域的直径进行了测量,PA0012和PA4324突变后,运动区域直径显著增加且能够被互补(图3B~C)。结果表明,PA0012和PA4324对铜绿假单胞菌的群集运动均具有显著的负调控作用。
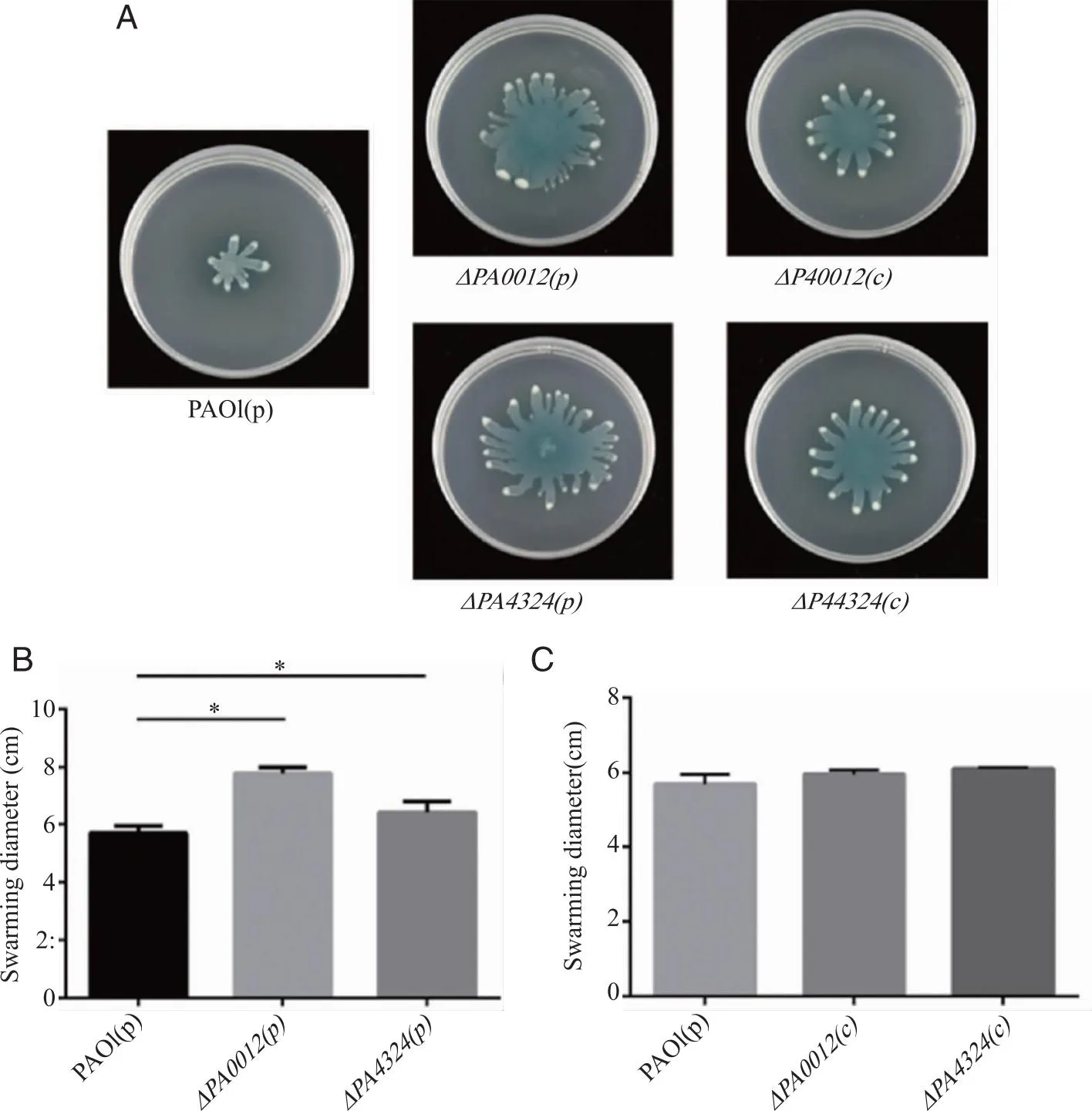
A:2个含有空质粒的单PilZ结构域蛋白突变体ΔPA0012 (p), ΔPA4324 (p)及其互补体ΔPA0012 (c), ΔPA4324 (c)与含有空质粒的野生型菌株PAO1 (p)群集运动能力的比较;B:2个含有空质粒的单PilZ结构域蛋白突变体ΔPA0012 (p)、 ΔPA4324 (p)与含有空质粒的野生型菌株PAO1 (p)群集运动区域直径的比较;C:2个单PilZ结构域蛋白突变体的互补体ΔPA0012(c)、 ΔPA4324 (c)与含有空质粒的野生型菌株PAO1 (p)群集运动区域直径的比较。图3 群集运动表型
2.4ΔPA0012和ΔPA4324慢杀死秀丽隐杆线虫的能力和蛋白酶活性无显著改变 秀丽隐杆线虫是一种模式动物,相关研究中用它作为寄主,模拟铜绿假单胞菌对人或者动物的感染。本研究用ΔPA0012,ΔPA4324和野生型菌株PAO1分别对线虫进行了慢杀死实验,结果显示ΔPA0012和ΔPA4324均与野生型菌株无显著性差异(P>0.05,图4A)。本实验以大肠杆菌OP50作为阴性对照。上述结果表明,PA0012和PA4324基因突变均不会显著影响铜绿假单胞菌PAO1对秀丽隐杆线虫的慢杀死。
蛋白酶包括多个种类,它们的活性在铜绿假单胞菌感染寄主时能够起到重要作用,它们能够水解寄主的免疫蛋白,从而导致发病。本实验用脱脂牛奶固体培养基检测了ΔPA0012、ΔPA4324和野生型菌株PAO1产生的总蛋白酶的水解活性。实验结果显示,相比较于野生菌株,2个突变菌株产生的蛋白水解酶的活性没有显著性改变(图4B)。该结果表明PA0012和PA4324基因突变均不会显著影响铜绿假单胞菌PAO1总蛋白酶对脱脂奶粉的水解活性。
2.5 ΔPA0012和ΔPA4324感染生菜的能力显著减轻 铜绿假单胞菌能够感染十字花科植物早已经被报道过。本研究用ΔPA0012和ΔPA4324分别去感染生菜,在每片生菜叶上同样也接种野生型菌株PAO1,比较同一片菜叶上突变体菌株与野生型菌株感染的差别,所有细菌均用硫酸镁溶液悬浮,并且以硫酸镁溶液作为对照,以防止叶片因缺镁产生变化。本研究的实验结果显示,相比较于野生型菌株PAO1,2个突变菌株感染生菜的情况显著减轻(图5)。该实验结果表明,PA0012和PA4324基因突变均能导致铜绿假单胞菌感染生菜的情况显著减轻。

A:突变体ΔPA0012与野生型菌株PAO1感染生菜的比较;B:突变体ΔPA4324与野生型菌株PAO1感染生菜的比较。图5 ΔPA0012和ΔPA4324感染生菜表型
2.6 ΔPA0012和ΔPA4324的RNA测序分析 在进行了表型筛选之后,进行了RNA测序分析。本研究以log2(基因表达倍数变化) ≥ 0或 ≤ 0为界限,结果显示,相比较于野生型菌株PAO1,ΔPA0012有612个基因表达量上调,有144个基因表达量下调;ΔPA4324有521个基因表达量上调,有102个基因表达量下调(图6A)。ΔPA0012所引起表达量有差异的基因广泛分布于微生物代谢等在内的多种通路中,将这些差异基因的功能分类,又可分为代谢、催化等多种类型,如其通路富集统计散点图(前20位)所示(图6B)。 ΔPA4324所引起表达量有差异的基因广泛分布于包括核糖体等在内的多种通路中,如其通路富集统计散点图(前20位)所示(图6C)。将这些差异基因的功能分类,又可分为代谢、结合和催化等多种类型。本研究用韦恩图对2个突变体的共有差异基因进行了分析,发现相比较于野生型菌株,ΔPA0012与ΔPA4324有148个共有差异基因(图6D)。
以上RNA测序结果表明,2个突变体有一些共同的差异基因,暗示了它们在调控铜绿假单胞菌生理进程的过程中可能存在着某些相近的方式。然而,PA0012和PA4324基因的突变也导致了铜绿假单胞菌中某些不同基因的差异表达。PA0012和PA4324这2个蛋白对铜绿假单胞菌生理进程的调控具有复杂的网络,还需进一步结合它们的互作蛋白进行研究。
3 讨论
本文对铜绿假单胞菌PA0012和PA4324这2个单PilZ结构域蛋白突变体的主要生物学表型进行了初步筛选,筛选结果表明,虽然它们影响的生物学表型较为相似,但转录组分析还是有明显的差异。根据蛋白氨基酸序列的差异,自然界中的单PilZ结构域蛋白又可以分为很多类,因此不同氨基酸序列的单PilZ结构域蛋白可能具有不同的生物学功能。本次研究的这2个蛋白氨基酸序列存在差异,说明它们的转录组存在差异也是合理的。虽然RNA测序的结果还需要进一步验证,但是,这些结果结合前期表型筛选的结果已经足以表明PA0012和PA4324能够影响铜绿假单胞菌的生物学表型。
根据本研究的实验结果,PA0012和PA4324的突变均能够导致铜绿假单胞菌PAO1的群集运动能力增强,并且可以被互补。C-di-GMP对群集运动的调控具有关键的作用,细胞内高浓度的c-di-GMP能够促进细菌生物被膜的形成,并且抑制其群集运动[11]。从2005年开始,研究人员就陆续发现一些能够调控生物被膜形成的c-di-GMP代谢酶,随着研究的不断深入,近几年又发现RmcA、WspR、MorA、PipA、SiaD和SiaA等c-di-GMP的代谢酶也能够调控绿脓杆菌生物被膜的形成[12-16],也有研究者发现c-di-GMP受体蛋白FlgZ能够调节铜绿假单胞菌的群集运动[17],FlgZ 蛋白是含有PilZ结构域的双结构域蛋白,根据报道,它能够依赖于c-di-GMP与运动促进定子MotC特异地互作,从而影响细菌的群集运动[17]。在铜绿假单胞菌编码的4个可结合c-di-GMP的单PilZ结构域蛋白中,前期报道了HapZ能够与杂合组氨酸激酶SagS相互作用[7],MapZ能跟甲基转移酶CheR1相互作用[8],而对于PA0012和PA4324这2个蛋白,了解还很有限。
C-di-GMP通路能够影响多种植物和人体/动物病原菌的毒性。C-di-GMP信号通路首次被发现能够影响毒性是在霍乱弧菌中(V.cholerae),高c-di-GMP浓度能够减轻霍乱弧菌对幼鼠的毒性。通过之前的研究,人们认为细菌的急性感染只需要低浓度的c-di-GMP,甚至不需要c-di-GMP,比如将鼠疫耶尔森菌(Yersiniapestis)中的某些有功能的DGC敲除以后并不影响细菌对小鼠的毒性[18]。有研究者将沙门菌(Salmonellaenterica)中所有含GGDEF结构域的基因敲除以后,发现细菌的毒性减弱,然而在突变体中互补一个没有c-di-GMP合成酶活性的二鸟苷酸环化酶STM4551就能将细菌对小鼠的毒性恢复回去,该结果表明c-di-GMP在沙门菌该种感染模型中不起作用[19]。还有研究者将羊布鲁杆菌(Brucellamelitensis)中所有的PDE进行了敲除,发现细菌感染小鼠的毒性会降低,而敲除所有的DGC之后则细菌对小鼠的毒性增强[20]。关于c-di-GMP代谢调控细菌毒性的报道还有很多,然而c-di-GMP受体PilZ结构域蛋白调控细菌毒性的报道还很有限。
C-di-GMP在许多环境或者病原微生物生理过程中都扮演了重要的角色,人们对其研究最多的是它在生物被膜形成和散播过程中所起的作用。本研究对铜绿假单胞菌PA0012和PA4324突变体生物学表型的筛选以及RNA测序分析让我们面对细菌致病过程中复杂的c-di-GMP受体蛋白信号通路有了一个新的视角,但同时也产生了一些疑惑,导致这些表型变化的机制是否为c-di-GMP在细胞内发挥作用还不得而知。本文筛选出了PA0012和PA4324能够显著影响铜绿假单胞菌的生物学表型,同时,推测了PA0012和PA4324在转录水平上可能参与调控细胞生理进程的通路,为更深入的理解和完善c-di-GMP信号传导系统对细菌致病性的调控作用提供了理论基础。
