CDC42 基因在成人和脐带血网织红细胞中差异表达的潜在价值
2023-12-02王进朝张根豪安阳市人民医院检验科河南安阳455000郑州大学第一附属医院输血科
王进朝 张根豪( 安阳市人民医院 检验科, 河南 安阳 455000;2 郑州大学第一附属医院 输血科)
镰状细胞病(sickle cell disease, SCD)是1 种β 球蛋白隐性突变导致镰状血红蛋白产生的常染色体疾病,严重时可危及患者生命[1]。 虽然近年来已有多项临床试验改善了SCD 患者的临床症状,但治疗效果仍不理想[2]。 胎儿血红蛋白(fetal hemoglobin, HbF)可以通过提高循环红细胞(red blood cells, RBC)的存活率改善SCD 患者的病情,这对探索新的SCD 治疗方法意义重大[3-5]。 血红蛋白成分在出生后会从HbF(α2γ2)转变为成人血红蛋白(adult hemoglobin,HbA, α2β2),这种现象称为血红蛋白转换[6-7]。 HbF 在人血红蛋白总量中所占比例极少,不到2%。 虽然已有研究报道高海拔或心肺疾病引起的缺氧可以诱导出生后γ-球蛋白和HbF 产生[8-9],但相关机制仍未清晰。
在本研究中,我们通过对GSE6236 和GSE17639 数据集中脐带血和成人外周血的网织红细胞的转录组进行全面分析,筛选出可能参与血红蛋白转换的差异基因,并在GSE35102 数据集、临床成人外周血和脐带血标本中进行验证,进而探索血红蛋白转换过程中的关键调控基因,为SCD患者的靶向治疗提供新的思路。
1 材料与方法
1.1 公共数据集下载和差异基因分析 从基因表达综合数据库(gene expression omnibus, GEO)下载GSE6236 (14 例脐带血,14 例成人外周血)、GSE17639 (6 例脐带血,6 例成人外周血)和GSE35102 中网织红细胞的转录组数据并使用Anno-Probe 包进行基因注释。 随后以|logFC|≥1.5 且P值<0.05 为阈值筛选差异基因。 GSE6236 和GSE17639 的转录组数据合并后使用Limma 包的removeBatchEffect 函数去除批次效应。
1.2 加权基因共表达网络分析 通过聚类树分析剔除离群值标本后,使用加权基因共表达网络分析(weighted gene coexpression network analysis, WGCNA)将基因聚类成不同颜色的模块,并分析模块与网织红细胞之间的相关性。 相关性系数>0.5 且P<0.001 的模块被认为是网织红细胞相关模块。
1.3 差异基因在血红蛋白转变过程中的表达量变化GSE35102 数据集是1 个将人原代红系祖细胞培养后进行基因表达谱分析的芯片数据。 研究人员将原代红系祖细胞在含有青霉素(100 U/mL)、链霉素(0.1 mg/mL)和30%胎牛血清的α-MEM 培养基内进行培养。 d0 在培养瓶内添加干细胞因子(50 ng/mL),白介素3(10 ng/mL)和促红细胞生成素(4 U/mL)等细胞因子。 分别在培养后的d7、14、21 和28 收集细胞并进行微阵列分析。 我们按照1、2、3 和4 周的时间间隔,将GSE35102 数据集中的标本分为4 组,用于验证差异基因在血红蛋白转变过程中的表达量变化。
1.4 实时定量PCR 检测差异基因在脐带血和成人外周血网织红细胞中的表达量 本研究获得了安阳市人民医院医学伦理委员会的批准(KS-2023-05-30)。 我们于2023 年6 月1 日—7月10 日分别从30 名成年健康志愿者和30 名产妇身上采集5mL外周血或脐带血。 如前所述[10-11],通过离心和过滤去除血浆、血小板和白细胞,获得富集的网织细胞。 使用TRIzol 试剂(Invitrogen,15596026)从富集的网织红细胞中分离总RNA,随后使用逆转录试剂将RNA 逆转录成cDNA(上海生工,B532441),最后按照说明书在Applied BiosystemsTM7500 仪器上进行实时PCR 反应。 CDC42 基因的引物序列为:上游CCATCGGAATATGTACCGACTG,下游CTCAGCGGTCGTAATCTGTCA; 内参基因β-ACTIN 的引物序列为:上游CGTGGGCCGCCCTAGGCACCA,下游TTGGCTTAGGGTTCAGGGGGG。
1.5 统计学分析 使用R.4.2 软件进行数据分析;使用独立样本t检验和Wilcoxon 秩合检验分析定量数据。 当P<0.05时认为结果具有统计学意义。
2 结果
2.1 差异基因筛选 如图1A 所示,在3 个数据集中,我们分别得到184,92,123 个差异基因(均|logFC|≥1.5 且P<0.05)。这些差异基因在脐带血和成人外周血网织红细胞中的表达如图1B 所示。 其中,12 个基因被认为是共同差异基因(图1C)。

图1 差异基因
2.2 数据集合并与批次效应去除 为了进一步分析脐带血与成人外周血中网织红细胞的转录组差异,我们将3 个数据集进行了合并。 合并后,共得到34 例脐带血网织红细胞标本和34 例成人外周血网织红细胞标本。 如图2A 所示,我们发现合并后的数据集具有明显的批次效应。 随后,我们使用Limma 包的removeBatchEffect 函数去除批次效应(图2B)。
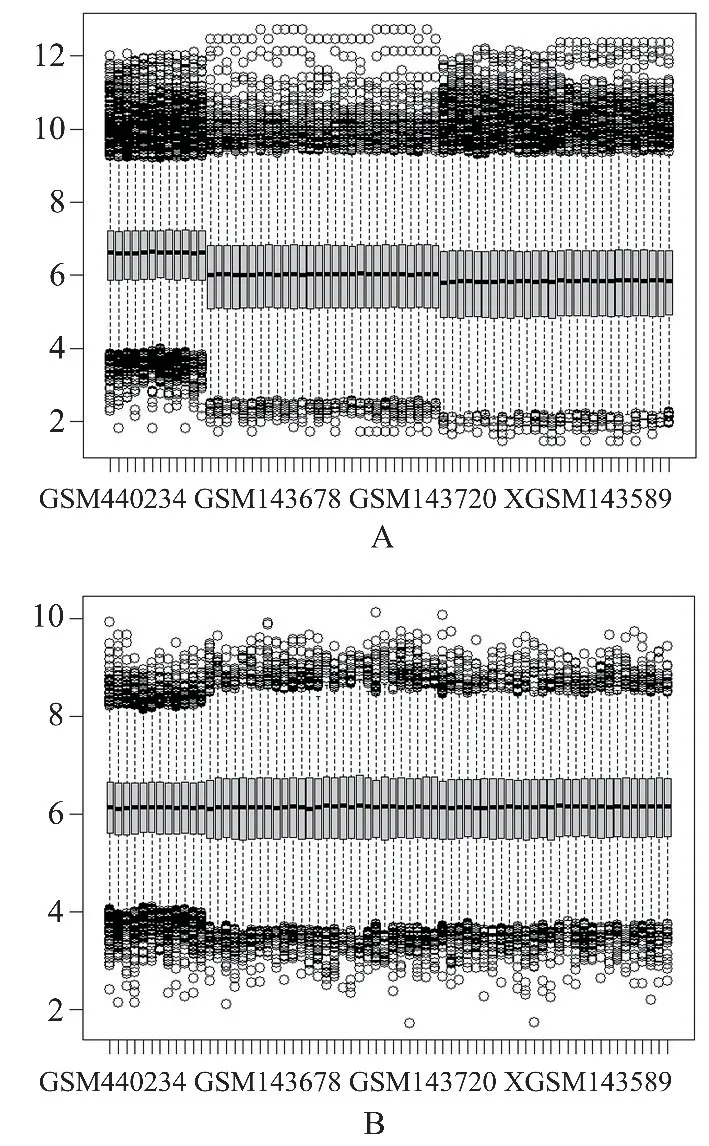
图2 批次效应去除
2.3 WGCNA 分析结果 我们首先对68 例标本进行聚类树分析,发现7 个离群值标本(图3A)。 将离群值标本剔除后,WGCNA 将基因聚类成蓝色和孔雀绿2 个不同颜色的模块(图3B)。 蓝色模块与网织红细胞相关性最大(相关系数=0.76,P<0.001),被认为是网织红细胞相关模块(图3C)。 蓝色模块共包含了718 个基因。
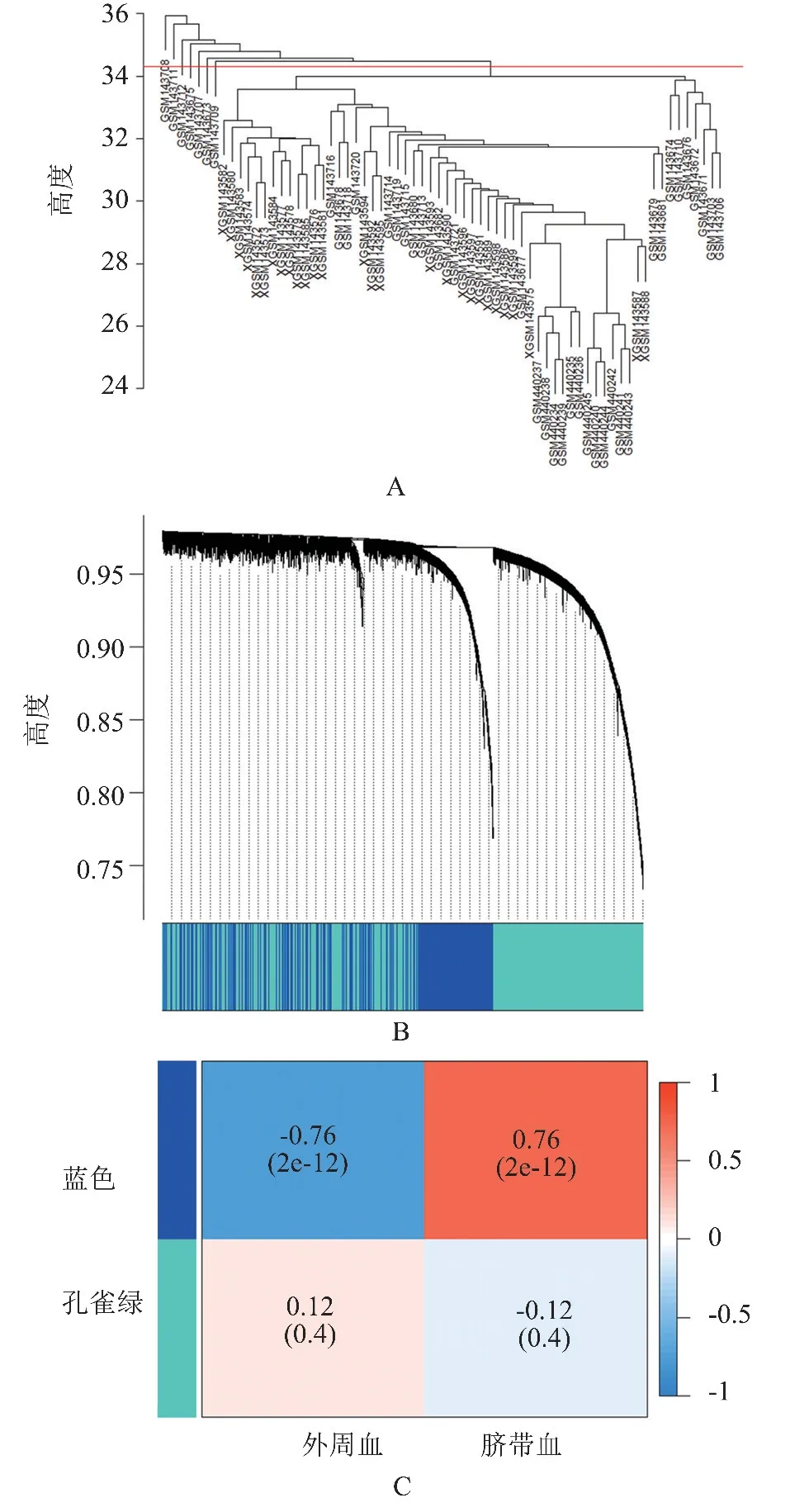
图3 WGCNA 分析
2.4CDC42 基因在脐带血和成人外周血网织红细胞中的差异性表达 如图4A 所示,我们将差异基因与蓝色模块的基因取交集后,共得到5 个基因。 然而在合并的数据集中,只有CDC42 基因的表达量具有差异性(t=3.776,P<0.001,图4B)。CDC42 基因可以很好的区分合并集中脐带血和成人外周血中的网织红细胞,曲线下面积(area under the curve, AUC)为0.747(图4C)。
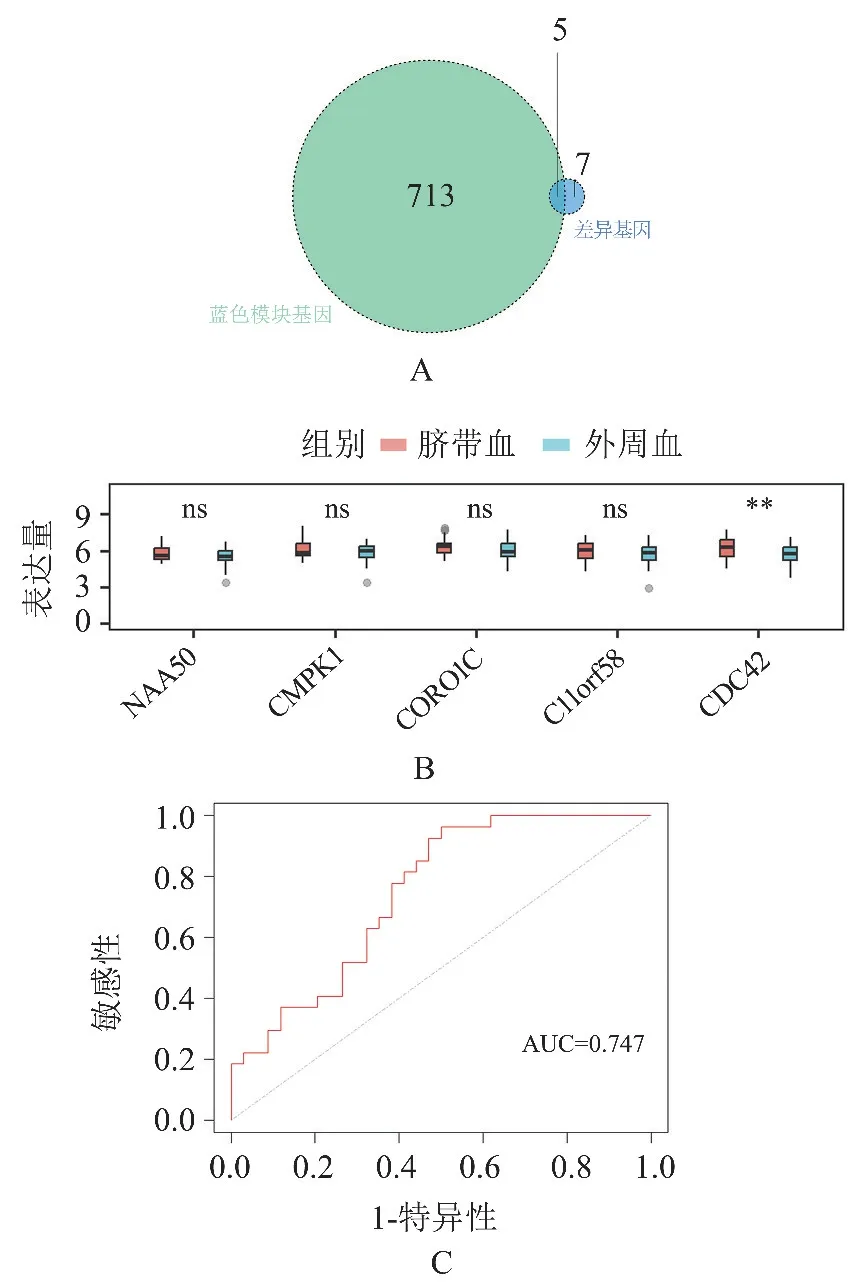
图4 CDC42 基因在合并数据集中的表达
2.5CDC42 基因在血红蛋白转变过程中的表达量变化 如图5A 所示,原代红系祖细胞培养后γ-球蛋白含量逐渐降低,而β-球蛋白含量逐渐升高,且二者在培养后第3 周左右出现了交集,表明此时发生了HbF/HbA 血红蛋白转变。 而CDC42 基因的表达量在培养后第2 周和第3 周的细胞中出现差异性改变(Z=-2.908,P<0.01,图5B),证明CDC42 基因可能参与了HbF/HbA 血红蛋白转变的进程。
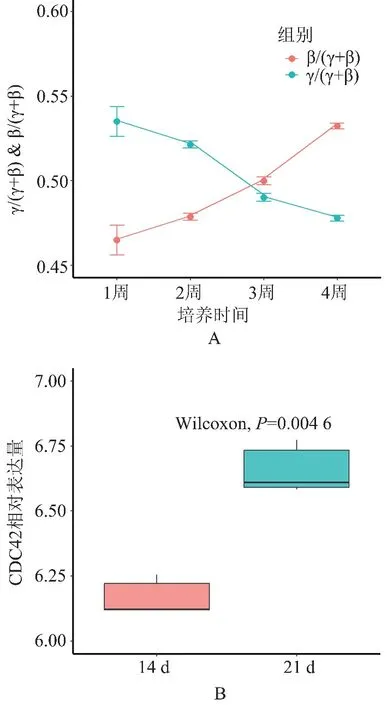
图5 CDC42 基因表达量在血红蛋白转变过程中的变化
2.6CDC42 基因在脐带血和成人外周血网织红细胞中表达量的差异分析 如图6A 所示,与脐带血中的网织红细胞相比(4.33±1.21),成人网织红细胞(2.47±0.85)中CDC42 基因的表达量显著降低(t=7.824,P<0.001),且CDC42 基因可以很好的区分脐带血和成人外周血中的网织红细胞,曲线下面积为0.892(图6B)。

图6 CDC42 基因在脐带血和成人外周血网织红细胞中表达量分析
3 讨论
SCD 是1 种血红蛋白β-珠蛋白链发生遗传性突变的血红蛋白病,其特征是慢性溶血性贫血、严重的急慢性疼痛以及伴随终身的末端器官损伤。 SCD 与早期死亡率密切相关[12]。 早期诊断、预防并发症和处理末端器官损伤等对症治疗对于改善SCD 的预后意义重大。 尽管美国食品和药物管理局已经批准了除羟基脲以外的3 种治疗SCD 的药物(Lglutamine, crizanlizumab 和voxelotor),但它们仍然不能从根本上治愈SCD[2]。 因此,临床工作中迫切地需要1 种更有效的方法改善SCD 患者的预后。
在本次研究中,我们通过对GSE6236 和GSE17639 数据集中脐带血和成人外周血中网织红细胞的转录组进行全面分析,筛选出5 个可能参与血红蛋白转换的差异基因。 虽然只有CDC42 基因在去除批次效应的合并数据集中有显著表达差异,但其可以很好地区分脐带血和成人外周血中的网织红细胞(AUC=0.747)。 随后,我们在GSE35102 数据集中进行验证。 GSE35102 数据集对原代红系祖细胞进行了全局基因表达谱分析,以探索在整个体外红系分化过程中参与γ/β-球蛋白转换的基因表达谱。 我们观察到原代红系祖细胞培养后γ-球蛋白含量逐渐降低,而β-球蛋白含量逐渐升高,且二者在培养后第3 周左右出现了交集,表明此时发生了HbF/HbA 血红蛋白转变。 而CDC42 基因的表达量在培养后第2 周和第3 周出现差异性改变,表明CDC42 基因可能参与了HbF/HbA 血红蛋白转变的进程。 至于其调节血红蛋白转变进程的具体分子机制,需要在未来的工作中进一步研究。
CDC42 基因是鸟苷三磷酸酶(GTPase)家族的1 个重要成员,参与调节细胞受到细胞外刺激后胞内三磷酸鸟苷(GTP)和二磷酸鸟苷的活性状态。CDC42 基因不但参与调节造血干细胞的粘附、迁移、归巢以及细胞周期等生物进程,在红细胞的最终分化过程中也起着至关重要的作用[13]。CDC42 基因缺陷小鼠会出现致命的骨髓增生性疾病,主要表现为髓系和红系发育抑制,以及显著的白细胞增多伴中性粒细胞增多[14]。 本研究发现CDC42 基因在脐带血网织红细胞中的表达显著高于成人外周血网织红细胞中的表达,且在原代红系祖细胞的γ/β-球蛋白转换过程中也存在差异性改变。HbF/HbA 血红蛋白的转变通常发生在婴儿出生后几个月内。 HbF 的生成增加可显著改善成人SCD 患者的临床症状。 目前通过靶向HbF/HbA 血红蛋白转变治疗SCD 患者的基因疗法主要包括2 种,1 种是通过慢病毒载体向红细胞内添加正常的β-球蛋白编码序列以增加HbF 的表达[15],另1种是通过Cas9 特异性敲除红细胞内的BCL11A 以重新激活γ-球蛋白基因[16-17]。 但这2 种方法的终末有效性和安全性仍值得商榷[18]。 考虑到CDC42 基因信号传导贯穿整个红细胞发育成熟的进程中[14],且CDC42 基因可能参与了HbF/HbA 血红蛋白转变进程,我们或许可以将CDC42 基因作为1个潜在的治疗靶点应用于SCD 患者的基因治疗。
综上所述,我们通过对脐带血和成人外周血中网织红细胞转录组的分析,发现CDC42 基因可能参与了HbF/HbA 血红蛋白转变的进程,可作为未来SCD 患者的潜在治疗靶点。在明确CDC42 基因参与HbF/HbA 血红蛋白转变进程的具体分子机制后,通过质粒或慢病毒转染的方式在网织红细胞内过表达CDC42 基因从而促进红细胞内HbF 表达增加,或许可以作为未来SCD 患者靶向HbF/HbA 血红蛋白转变的潜在治疗靶点,为SCD 的基因疗法开辟1 种新的方向。
利益冲突:所有作者均声明不存在利益冲突。
