动物内寄生虫丝氨酸蛋白酶研究进展
2023-10-24陈奕君
陈奕君,周 璇,谢 跃
(四川农业大学动物医学院,四川成都 611130)
丝氨酸蛋白酶(serine proteases)是一类存在于真核生物、原核生物和病毒的蛋白水解酶,该酶因活性位点丝氨酸残基对底物的亲核攻击而得名丝氨酸蛋白酶。与金属、半胱氨酸和天冬氨酸蛋白酶类似,丝氨酸蛋白酶通过分裂肽键进行蛋白质水解反应[1],广泛参与动物机体的蛋白质代谢、消化、凝血、发育调节和受精等生理过程,并与许多人类疾病的发生息息相关[2]。内寄生虫作为人和家养及野生动物常见的一类病原,可引起动物贫血、厌食、消瘦、衰弱、甚至导致死亡[3]。内寄生虫丝氨酸蛋白酶是虫体感染宿主的关键蛋白分子,参与虫体的发育和营养摄取以及对宿主入侵、抗凝和免疫逃逸等生物学过程[4]。肠道蠕虫丝氨酸蛋白酶的研究证实,该酶可参与繁殖、躲避宿主免疫系统和发育等生理过程。鉴于内寄生虫丝氨酸蛋白酶重要的分子特性及其在寄生虫-宿主互作中的关键作用,它被视为新一代抗寄生虫干预免疫治疗、化疗以及血清诊断试剂的重要靶点。有关动物内寄生虫丝氨酸蛋白酶的研究较零散,缺乏系统性归纳和总结,本综述拟对动物内寄生虫如原虫、绦虫、吸虫和线虫的丝氨酸蛋白酶研究现状及其在寄生虫-宿主互作中的生物学功能进行概述,以期为该蛋白酶的进一步研究提供信息参考。
1 丝氨酸蛋白酶概述
丝氨酸蛋白酶主要水解蛋白肽键,其结构由β蛋白构成,且包含两个相似结构域,即N结构域和C结构域。丝氨酸蛋白酶的活性催化位点及其重要的功能位点均位于N结构域和C结构域,其中活性催化中心由来自N结构域的两个残基(His/Asp/Lys/Glu)以及C结构域的Ser残基共同构成,而其他功能位点包括氧离子孔、底物特异性口袋和底物结合位点的残基则相对可变,位于C结构域[5]。由此,在整个丝氨酸蛋白酶超家族成员中,有些丝氨酸蛋白酶氨基酸序列间的相似性甚至不到20%,这些序列和结构差异,导致了丝氨酸蛋白酶功能上的变化,给整个超家族的遗传进化与功能研究带来了一定困难[6]。目前,丝氨酸蛋白酶主要通过底物的特异性以及催化位点的拓扑结构来进行分类。例如,根据底物的特异性,丝氨酸蛋白酶可分为胰蛋白酶样、凝乳蛋白酶样、枯草蛋白酶样、弹性蛋白酶样和激肽酶[5];根据催化位点的拓扑结构,丝氨酸蛋白酶可分为13个家族和40科[6](表1)。基于储存在MEROPS数据库数据,PA是丝氨酸蛋白酶超家族中最大的家族。该家族主要在真核生物中表达,而在原核生物或植物中的表达却极少[3]。
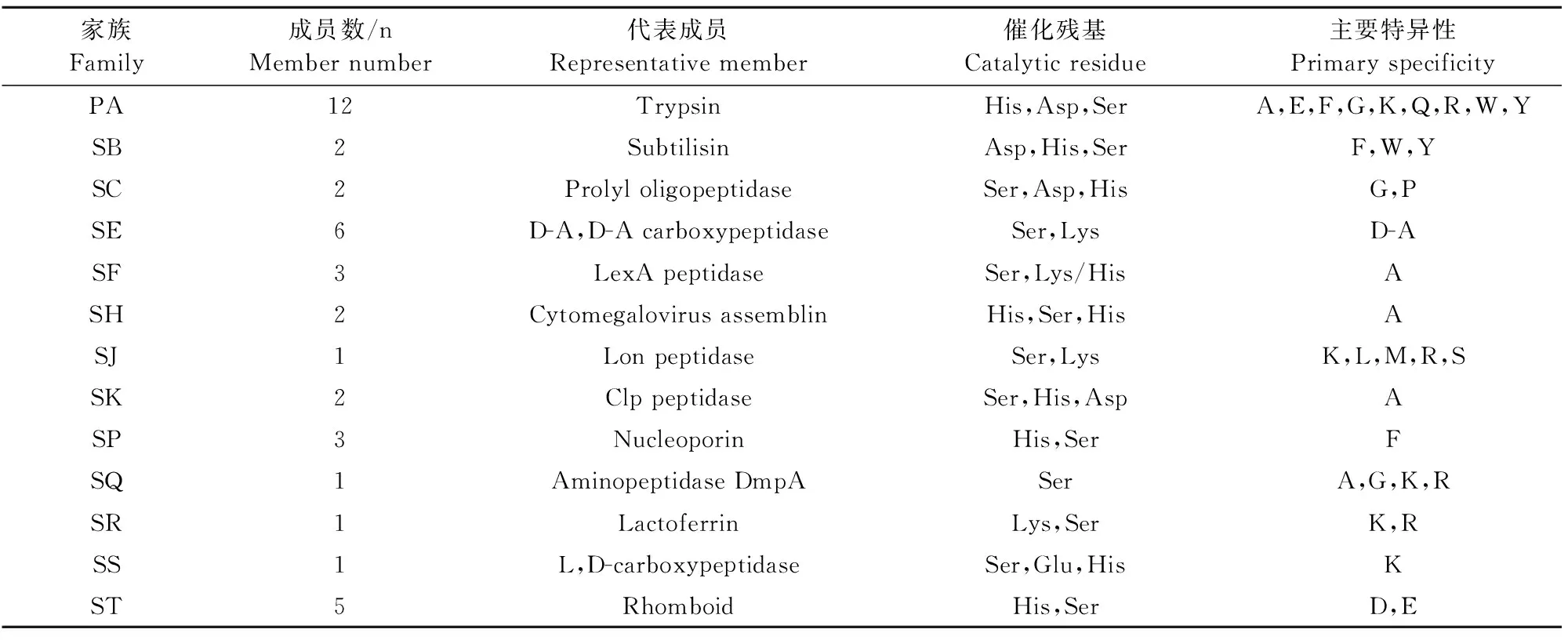
表1 丝氨酸蛋白酶家族Table 1 The family of serine proteases
2 动物内寄生虫丝氨酸蛋白酶
动物内寄生虫是一类寄生于人和动物体内的寄生虫,主要包括原虫、吸虫、绦虫及线虫。这些寄生虫通过抢夺宿主营养物质、破坏宿主组织、释放有毒物质(如排泄分泌抗原)等行为,引起宿主动物出现营养不良、贫血,组织器官病变,过敏反应,甚至死亡[3]。研究表明,丝氨酸蛋白酶作为内寄生虫重要的分泌排泄蛋白,在虫体的宿主入侵过程中扮演着重要角色,是内寄生虫建立有效感染的关键蛋白[4]。例如,恶性疟原虫丝氨酸蛋白酶PfSUB1被证实与虫体入侵宿主红细胞有关[7],利什曼原虫丝氨酸蛋白酶LaSP-Sol被证实参与介导宿主免疫反应[8],刚地弓形虫菱形蛋白酶更是被证实与虫体在宿主组织中的转移有关[9]。在动物吸虫中,肝片吸虫的丝氨酸蛋白酶被证实具有水解蛋白和促进虫体营养的作用[6],血吸虫的丝氨酸蛋白酶SmSPs1-5更是被证实具有抗凝和调节宿主血压的作用[10]。在动物绦虫、线虫中,丝氨酸蛋白酶还被证实具有调控虫体发育[11]、参与虫体宿主寄生与繁殖的作用[12]。由此可见,动物内寄生虫丝氨酸蛋白酶在虫体寄生及其与宿主互作的过程中发挥着重要的作用。目前有关内寄生虫丝氨酸蛋白酶及其类别与功能如表2所示。
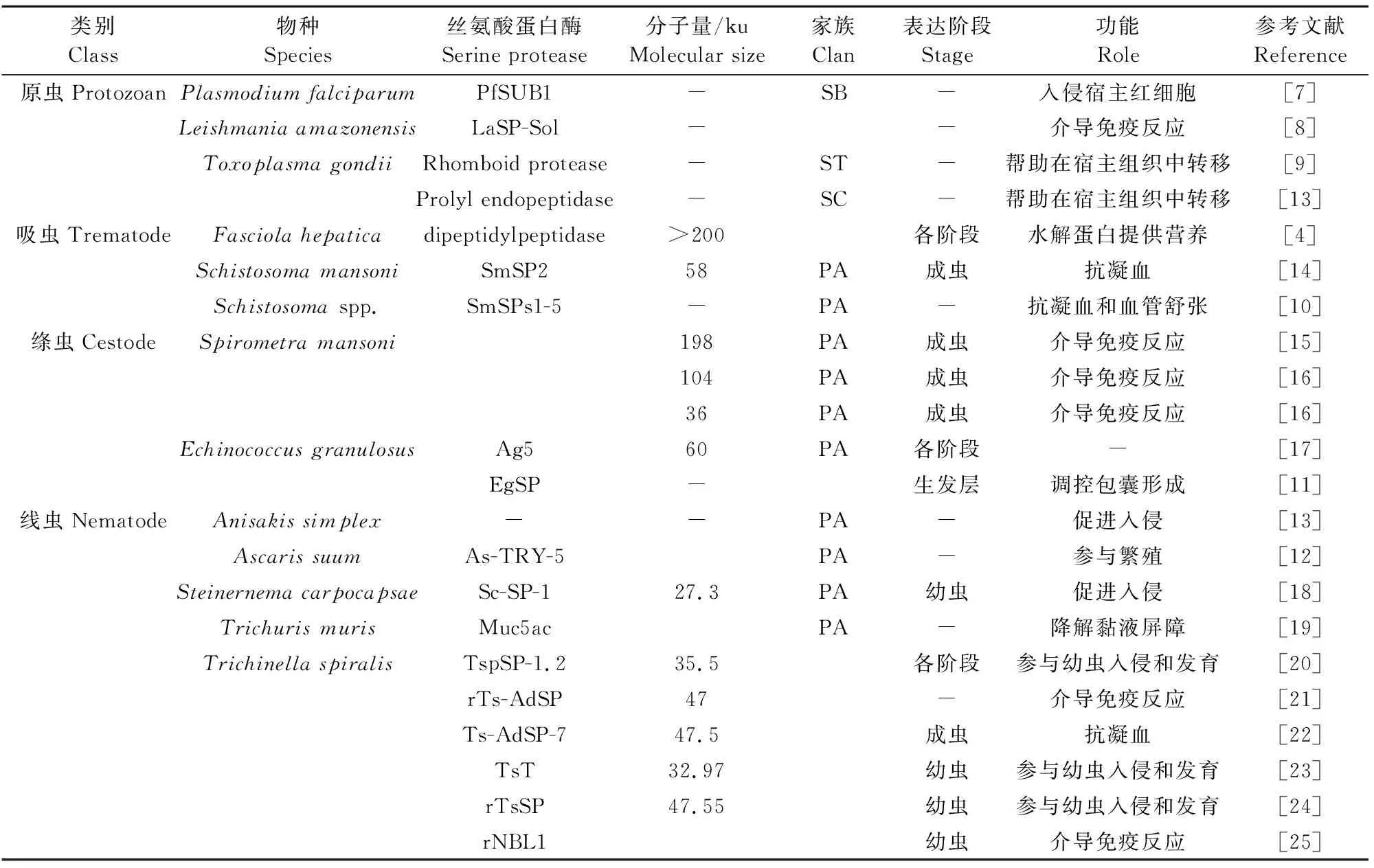
表2 内寄生虫丝氨酸蛋白酶的特征Table 2 Features of serine proteases in endoparasites
3 动物内寄生虫丝氨酸蛋白酶的功能
3.1 宿主入侵
目前发现原虫、线虫和吸虫的一些丝氨酸蛋白酶可有参与内寄生虫感染入侵宿主的过程。这些丝氨酸蛋白酶可以通过调控与入侵相关分子的释放或是降解消化宿主组织细胞从而促进虫体入侵宿主,其还可以促进虫体在宿主体内组织中的传播。在恶性疟原虫的相关研究中,研究人员发现疟原虫枯草杆菌素样丝氨酸蛋白酶(PfSUB1)在虫体裂殖子发育的最后阶段,可被释放到红细胞纳虫空泡(parasitophorous vacuole),对SERA家族木瓜蛋白酶类蛋白进行蛋白质修饰,从而有利于宿主细胞破裂后释放裂殖子,入侵新的红细胞。此外,PfSUB1还作为一种多功能加工蛋白酶,参与原虫虫体对宿主细胞表面的重塑,促进虫体在组织间的传播[7,26-27]。例如,刚地弓形虫丝氨酸蛋白酶通过重塑宿主细胞表面的结构,协助刚地弓形虫速殖子侵入上皮细胞及其在组织间的传播。同时研究证实,利什曼原虫丝氨酸蛋白酶SB族、SC族和SF族可以直接参与虫体对宿主的入侵过程[8,28]。在线虫丝氨酸蛋白酶的研究中,Sakanari J A等发现简单尖异线虫(Anisakissimplex)幼虫含有一种25 ku的胰蛋白酶样丝氨酸蛋白,可以降解结缔组织胞外基质,进而有利于虫体穿透宿主胃和肠的黏膜层、黏膜下层和肌层,向网膜、肝脏、胰腺或胆囊迁移[13],与此同时,鼠鞭虫成虫分泌一种丝氨酸蛋白酶样物质可以通过破坏肠道黏液屏障以及上皮细胞膜的完整性(包括降解肠道黏蛋白Muc2)从而解开黏液网,促进虫体在入侵和肠道定植等过程,参与鼠鞭虫对宿主的致病作用[19]。此外,在斯氏线虫中发现的似丝氨酸蛋白酶样蛋白(Sc-SP-1)仅在虫体寄生阶段表达,并且Sc-SP-1在准备入侵血腔的虫体中表达量高于血腔中的已有线虫。同时,体外试验证实,Sc-SP-1能与昆虫宿主中肠上皮结合引起细胞脱落,并能够导致人造膜形成孔洞,因此Sc-SP-1的致病功能可能是通过破坏中肠屏障而侵入血腹腔实现[18]。既往研究表明,旋毛虫丝氨酸蛋白酶可破坏肠道上皮细胞膜的完整性,这是由于其水解宿主肠道表面的黏液屏障[20]。此外,相关学者发现,在虫体的整个生命周期中,肌肉移行期的旋毛虫幼虫的排泄-分泌蛋白(ES蛋白)与虫体提取物均表现出大量的丝氨酸蛋白酶活性,这种阶段特异性的蛋白水解活性有助于在寄生虫感染期间破坏宿主内部的机械和体液屏障。这些丝氨酸蛋白酶是抗体反应的靶点,可以抑制蛋白酶活性,并可能导致致敏宿主中寄生虫的损伤。有研究者利用RNAi技术沉默TsSP1.2基因后,发现与未干扰组相比,回收的新生幼虫数量以及各阶段虫体的长度都出现显著降低,说明TsSP1.2在虫体繁殖、入侵和发育过程中起着至关重要的作用[24,30]。在血吸虫的研究中,曼氏血吸虫发现存在一种具有凝乳蛋白酶样活性的弹性蛋白酶(SmCE),通过免疫定位证实虫体离开中间宿主前SmCE被激活,推测SmCE参与虫体皮肤侵袭过程[31]。该蛋白酶除了降解弹性蛋白外,还能降解结缔组织中的其他蛋白质(例如胶原蛋白、纤连蛋白和免疫球蛋白),并与蠕虫用来消化血液的半胱氨酸和天冬氨酸蛋白酶具有功能相似性[15]。
3.2 凝血和纤溶
3.3 其他
除上述功能之外,有研究表明丝氨酸蛋白酶参与内寄生虫的繁殖过程。如猪蛔虫丝氨酸蛋白酶在虫体繁殖交配过程中有促进精子活化样的作用[12]。另外,丝氨酸蛋白酶还参与内寄生虫虫体营养吸收过程,为寄生虫提供生长营养物质从而促进虫体生长发育[4]。在原虫中发现的丝氨酸蛋白酶可以影响原虫子孢子的活力及排出[33],还有研究推测细粒棘球绦虫中的丝氨酸蛋白与包囊的形成有关[11]。
4 动物内寄生虫丝氨酸蛋白酶的免疫保护抗原特性
丝氨酸蛋白酶作为一种分泌排泄蛋白,可以引起宿主免疫反应。在动物绦虫蚴病研究中,Kong Y等(1994)从曼氏裂头蚴中分离得到3种天然丝氨酸蛋白酶,经血清学试验证实,3种丝氨酸蛋白酶可与曼氏裂头蚴感染患者的血清IgG特异性结合,是裂头蚴病的潜在诊断抗原的有效候选[16]。另外,研究人员发现,人兽共患细粒棘球蚴可以分泌一种类丝氨酸蛋白酶AG5,且该蛋白在整个虫体生活史周期均有表达,因此推测该蛋白可能作为一种潜在的抗原,用于细粒棘球绦虫病中间宿主或最终宿主血清学诊断和疫苗开发[17]。类似的,在动物线虫病研究中,Liu M Y等筛选和鉴定到一种旋毛虫新生幼虫阶段特异性的丝氨酸蛋白酶NBL1[25]。结果分析揭示,该蛋白包括1个催化结构域和1个C端结构域。重组蛋白rNBL1的截断变体表位定位发现,NBL1 C端结构域为其免疫显性区域,具有较好的免疫原性。借助旋毛虫-猪感染模型,研究人员证实NBL1蛋白可以诱导宿主产生惊人的免疫保护力,抵抗旋毛虫早期感染[4]。另外,Feng S等对重组旋毛虫丝氨酸蛋白酶蛋白(rTs-Adsp)进行小鼠免疫保护试验,结果显示,重组蛋白rTs-Adsp可诱导小鼠产生较高水平的特异性抗体IgG和IgE以及Th1(IFN-γ和IL-2)和Th2(IL-4、IL-10和IL-13)细胞因子,同时相较对照组,重组蛋白rTs-Adsp可有效降低小鼠肌肉中移行的幼虫数量,减虫率达46.5%,说明可以作为旋毛虫病的潜在候选疫苗[23]。Xu D等使用旋毛虫-猪感染模型重复了上述免疫试验,结果证实rTs-Adsp同样可以诱导实验猪产生较好的旋毛虫病免疫保护效果[21]。此外,Zhang Y等对另一种旋毛虫丝氨酸蛋白酶TsT进行免疫保护作用评价。结果表明,重组蛋白rTsT免疫小鼠的血清IgG、IgG1/IgG2a均显著上升,同时伴随肠道IgA以及细胞因子IFN-γ和IL-4的升高;免疫保护效果比较发现,相较对照组,重组蛋白rTsT免疫组小鼠肠道的成虫减少33.17%,肌肉幼虫减少37.80%,提示rTsT可作为另一旋毛虫病的疫苗候选抗原[34,35]。
5 展望
内寄生虫作为一类严重危害畜牧业发展和人类健康的重要病原,具有重要的公共卫生和社会经济影响。考虑到目前内寄生虫耐药性问题正变得日益严重,寻求新的药物靶点以及其他疫苗的研究变得十分必要和迫切。越来越多的组学研究以及积累的实验证据表明,内寄生虫丝氨酸蛋白酶广泛参与内寄生虫的营养发育、宿主组织和细胞入侵、抗凝和免疫反应等生物学过程,说明丝氨酸蛋白酶有望成为抗寄生虫的新型分子靶点。目前部分研究证实内寄生虫丝氨酸蛋白酶具有作为新型疫苗的潜质,但是有关内寄生虫丝氨酸蛋白酶是否可以作为疫苗用于寄生虫病防控,仍需要大量的试验进行验证。相信随着人们对寄生虫丝氨酸蛋白酶研究的不断深入,所得结果势必将为寄生虫丝氨酸蛋白酶的应用以及开发其在抗寄生虫中作用(药物或疫苗)提供理论依据和基础。
