运动干预对肥胖大鼠睾丸自噬和凋亡的影响*
2023-07-19徐瑞董佳曹友祥胡慧夏秋王鹏银郁丽冯婉瑜盛蕾
徐瑞 董佳 曹友祥 胡慧 夏秋 王鹏银 郁丽冯婉瑜 盛蕾**
(1)南京体育学院运动健康学院,南京 210014;2)江苏省运动与健康工程协同创新中心,南京 210014;3)上海体育学院运动科学学院,上海 200438)
肥胖是一种由行为、环境和遗传原因引起的代谢紊乱疾病,除导致2型糖尿病、高血压、胰岛素抵抗等代谢紊乱疾病发生外,也会对生殖功能产生不良影响。男性不育症随着肥胖和代谢综合征的增加而持续增加[1-2]。肥胖会诱发精子质量下降、生殖细胞凋亡、精子DNA损伤、睾酮分泌不足等生殖功能相关问题的发生[3-5]。随研究的深入发现,高脂饮食诱导的肥胖[6]、糖尿病[7]等能量过剩条件可能通过自噬诱导精子缺陷和睾丸损伤的发生。自噬是依靠溶酶体的保守的自我消化过程,通过清除蛋白质聚集体、受损细胞器和脂滴起到维持细胞内稳态的保护作用[8],在睾丸组织中广泛参与精子发生过程,参与精子细胞顶体形成和鞭毛发育,去除不必要的细胞质,并通过改变细胞骨架重塑细胞形状[9]。特异性敲除生殖细胞中自噬相关基因ATG5、ATG7会导致小鼠出现精子数量降低,生育能力下降的现象[10-11]。当自噬不足以保护细胞免受应激损伤时,细胞就会发生凋亡,二者往往相伴出现。睾丸细胞中适当的凋亡能够清除异常及过量的生精细胞,以保证精子的正常发育和成熟,但过度的凋亡会导致少精或弱精[12-13]。
脂肪过度堆积导致的脂毒性损伤可能会抑制非脂肪组织的自噬[14]。肥胖和高脂饮食环境可能会通过抑制腺苷酸活化蛋白激酶(AMP-activated protein kinase,AMPK)磷酸化和激活哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)来抑制UNC-51样激酶1(ULK-51-like kinase 1,ULK1),从而抑制自噬的发生。也有研究发现,肥胖小鼠睾丸中自噬标志物微管相关轻链蛋白3(microtubule-associated protein 1-light chain 3,LC3)生成及选择性自噬接头蛋白1(sequestosome-1,SQSTM1/p62)降解减少[15]。提示肥胖可能抑制睾丸自噬过程的各个阶段。肥胖诱发自噬过程出现障碍的同时也会诱导生殖细胞凋亡[14]。肥胖会抑制抗凋亡的B淋巴细胞瘤2(B-cell lymphoma-2,BCL-2)蛋白,上调促凋亡的BCL-2相关X(BCL 2-associated X,Bax)蛋白和胱天蛋白酶(cysteinyl aspartate specific proteinase,Caspase)3表达,导致生殖细胞过度凋亡[16-18],并使睾丸组织中细胞数量减少[19],睾酮分泌随之受到抑制[20]。
AMPK作为一种能量感应激酶可被ATP消耗激活,其在激活自噬、抑制凋亡过程中发挥重要的生物学调节效应。一方面,磷酸化的AMPK可直接或间接磷酸化ULK1诱导自噬的启动[21]。另一方面,AMPK的激活已被证明可以防止高血糖或高脂肪诱导的血管内皮细胞和心肌细胞凋亡细胞死亡[22-23]。在睾丸组织中,AMPK抑制也被发现与睾丸间质细胞凋亡有关[24]。运动在促进能量消耗、改善肥胖对雄性生殖功能的负面影响,促进睾酮合成等方面的作用早被证实[25]。而在这一过程中,运动是否通过激活AMPK来逆转由于肥胖导致的睾丸自噬与凋亡异常,继而使睾丸的结构和功能得到改善?目前尚未可知。本研究通过高脂饮食喂养建立肥胖大鼠模型,并对其进行8周中等强度运动干预,观察运动和肥胖对大鼠生精小管结构及自噬和凋亡相关蛋白质的影响,旨在为运动改善肥胖群体睾丸组织损伤、生殖功能障碍的研究奠定理论基础。
1 材料与方法
1.1 实验动物及分组
清洁级6周龄SD雄性成年大鼠50只(初始体重为(220.6±5.3)g),购于浙江维通利华实验动物技术有限公司,生产许可证号为SMCXK(浙)2019-0001。适应性饲养1周后,将大鼠随机分为标准饲料(SLACOM,P1101F-25)饲养组(SD,n=20)和高脂饲料(Research Diet,D12451,能量配比为蛋白质20%、碳水化合物35%、脂肪45%)饲养组(HFD,n=30)。所有大鼠分笼饲养于江苏省农业科学院动物研究所鼠动物房(5只/笼),光照12 h/d,室温(24±4)℃,相对湿度(50±10)%,自由饮食。第8周末,将HFD组体重超过标准饮食组20%~30%(包含20%)者判定为肥胖大鼠(n=20)[26]。将SD组大鼠分为标准对照组(CC,n=10)和标准运动组(CE,n=10),将建模成功的肥胖大鼠分为肥胖对照组(OC,n=10),肥胖运动组(OE,n=10),所有大鼠维持原有饲养条件并对运动组大鼠施加运动干预。
本研究经南京体育学院实验动物伦理委员会批准,符合3R标准,批号为2019-009。
1.2 运动干预
CE和OE组大鼠进行为期8周的中等强度运动干预。正式运动前进行3 d适应性运动,15 min/d,适应性速度为 8 m/min。正式运动阶段进行60%~70% V·O2max强度的跑速,60 min/d,5 d/周。实验期间,运动速度参考Leandro等[27]改良测试V·O2max测试方案及本研究团队前期V·O2max测试结果[28]而定,每2周调整一次跑速(第1~2周v=11 m/min,第3~4周v=13 m/min,第5~6周v=14 m/min,第7~8周v=15 m/min)。
1.3 实验取材
运动干预第8周末,在最后一次运动48 h后,所有大鼠经20%乌拉坦溶液进行腹腔注射麻醉(1 ml/100 g体重),称量体重后腹主动脉取血,剥离睾丸并称重。根据体重和睾丸重量计算睾丸指数:睾丸指数=[(睾丸质量/g)/(身体质量/g)]×100%。其中一侧睾丸置于4%多聚甲醛固定液中,另侧睾丸在冰上剪碎分装至冻存管中,放入液氮中冷冻,取样结束后转移至-80℃冻存待用。
1.4 指标测试与方法
1.4.1 苏木精-伊红(HE)染色
将多聚甲醛固定的一侧睾丸石蜡包埋,进行横向切片,厚度为5 μm。石蜡切片经二甲苯脱蜡2次,乙醇梯度脱水处理后至苏木精溶液染色,染色后1%盐酸乙醇分化,再至伊红溶液染色,随后用乙醇溶液(50%、70%、90%、100%)分别冲洗5 min。二甲苯透明2次后,中性树胶封固。在显微镜100或200倍镜下每进行观察拍摄,每组随机选择5只大鼠,从每只大鼠中选取4个切片,利用显微镜选择视野内具有代表性的生精小管进行拍摄,采用ImageJ软件测量生精小管直径(单位:μm)及精原细胞个数[29]。共测量20个生精小管直径,并在20个下视野计数精原细胞数量。
1.4.2 蛋白质印迹法(Western blot)检测
取20 mg睾丸组织与400 μl加有蛋白酶抑制剂的裂解液于研磨管中,充分裂解,离心,取上清液。BCA法测定蛋白质浓度。配置浓度为10%和12.5%的SDS-PAGE胶,上样量8 μl,电泳恒压70 V,约60 min后转恒压90 V继续电泳75 min。湿转法PVDF膜转膜75 min后,5%脱脂牛奶封闭60 min,一抗按照AMPK(Abcam,ab32047,1∶6 000)、p62(Santa Cruz Biotechnology,sc-48402,1∶700)、GAPDH(CST,5174T,1∶1 000)、BCL-2(Proteintech,60178-1-lg,1∶8 000)、Bax(Proteintech,50599-2-lg,1∶10 000)、LC3(Proteintech,14600-1-AP,1∶2 000)的比例进行稀释,4℃摇床孵育过夜。次日,二抗室温孵育60 min,稀释比例为山羊抗小鼠(HUBIO,HA1006,1∶1 000),山羊抗兔(HUBIO,HA1006,1∶1 000)。化学发光凝胶成像仪暗室中曝光显影。采用ImageJ软件对蛋白质条带进行灰度值分析,以GAPDH为内参,目的蛋白相对表达量=目的蛋白灰度值/内参蛋白灰度值。
1.4.3 免疫荧光染色
石蜡切片脱蜡通透后至柠檬酸钠缓冲液进行抗原修复,PBS清洗3遍后滴加山羊血清,覆盖组织封闭30 min。随后进行一抗孵育,一抗稀释比例为BCL-2(Proteintech,60178-1-lg,1∶500)、LC3(Proteintech,14600-1-AP,1∶300),一抗4℃孵育过夜,次日复温PBS清洗,避光二抗孵育2 h,稀释比列为山羊抗兔(Abcam,ab150077,1∶500)、山羊抗小鼠(Abcam,ab150116,1∶500)稀释孵育2 h。最后滴加含DAPI的抗荧光猝灭剂,盖玻片轻盖封片。每组随机选择5张切片,在显微镜400倍镜下每张切片随机选择5个非连续视野进行观察拍摄。使用ImageJ软件对BCL-2和LC3免疫荧光进行定位和半定量图像分析。
1.5 数据统计分析
所得数据运用SPSS 22.0和R语言进行整理分析,采用Shapiro-Wilk法对各指标数值进行正态性检验,符合正态分布的数据以平均数±标准差()表示。以高脂饮食和运动作为影响因素进行2(干预前后)×2(高脂饮食和运动干预)双因素方差分析,对每个因素的主效应和交互作用进行Bonferroni检验;蛋白质指标间的相关关系表示为Pearson相关系数,相关系数0.8以上为极强相关,0.6~0.8为强相关,0.4~0.6为中等程度相关,0.2~0.4为弱相关,0.2以下为极弱相关或无相关。P<0.05为具有统计学意义。
2 结果
2.1 大鼠肥胖模型建立
大鼠基础体重为(220.6±5.3)g,分别对SD组和HFD组进行标准饲料和高脂饲料喂养。喂养2周后SD组大鼠体重为(349.79±15.3)g,HFD组大鼠体重为(376.19±14.13)g;喂养4周后SD组大鼠体重为(435.73±30.84)g,HFD组大鼠体重为(529.58±34.9)g;喂养6周后SD组大鼠体重为(487.49±28.68)g,HFD组大鼠体重为(573.06±51.29)g;喂养8周后SD组大鼠体重为(494.46±28.04)g,HFD组大鼠体重为(641.68±43.37)g(图1)。在8周建模结束后,HFD组的体重比SD组高29.77%(>20%),表明肥胖建模成功。

Fig. 1 The change of body mass during the establishment of obesity model

Fig. 2 Comparison of body mass and testis index in each group

Fig. 3 HE staining in each group
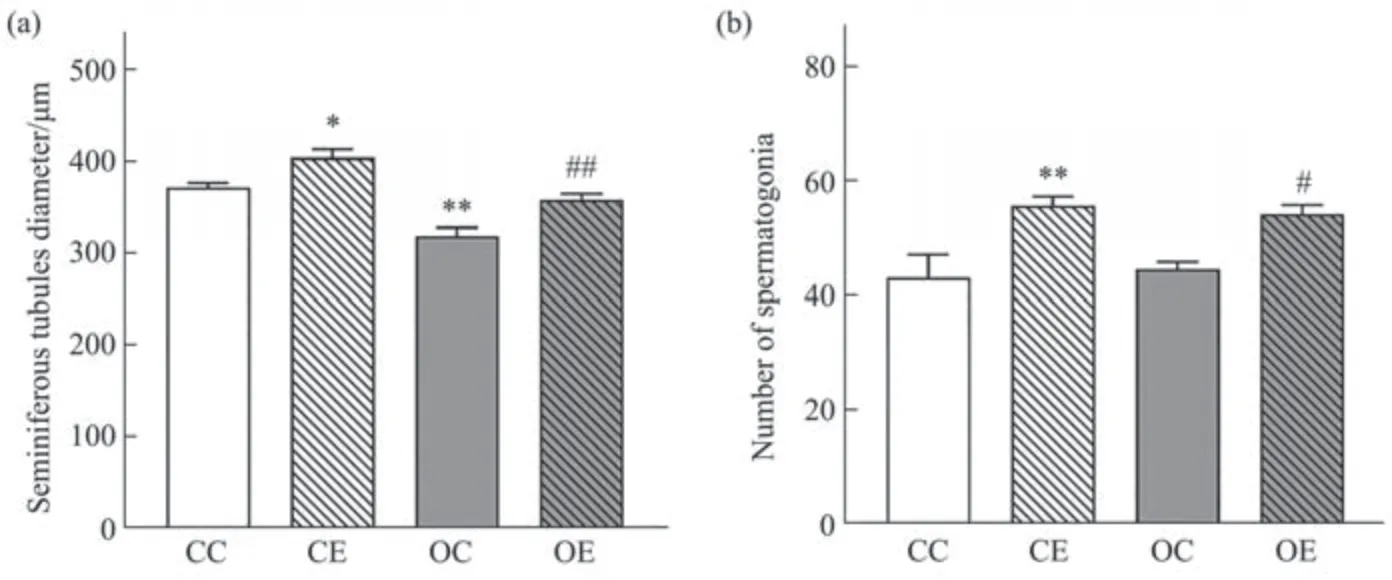
Fig. 4 Comparison of seminiferous tubules diameter and number of spermatogonia in each group
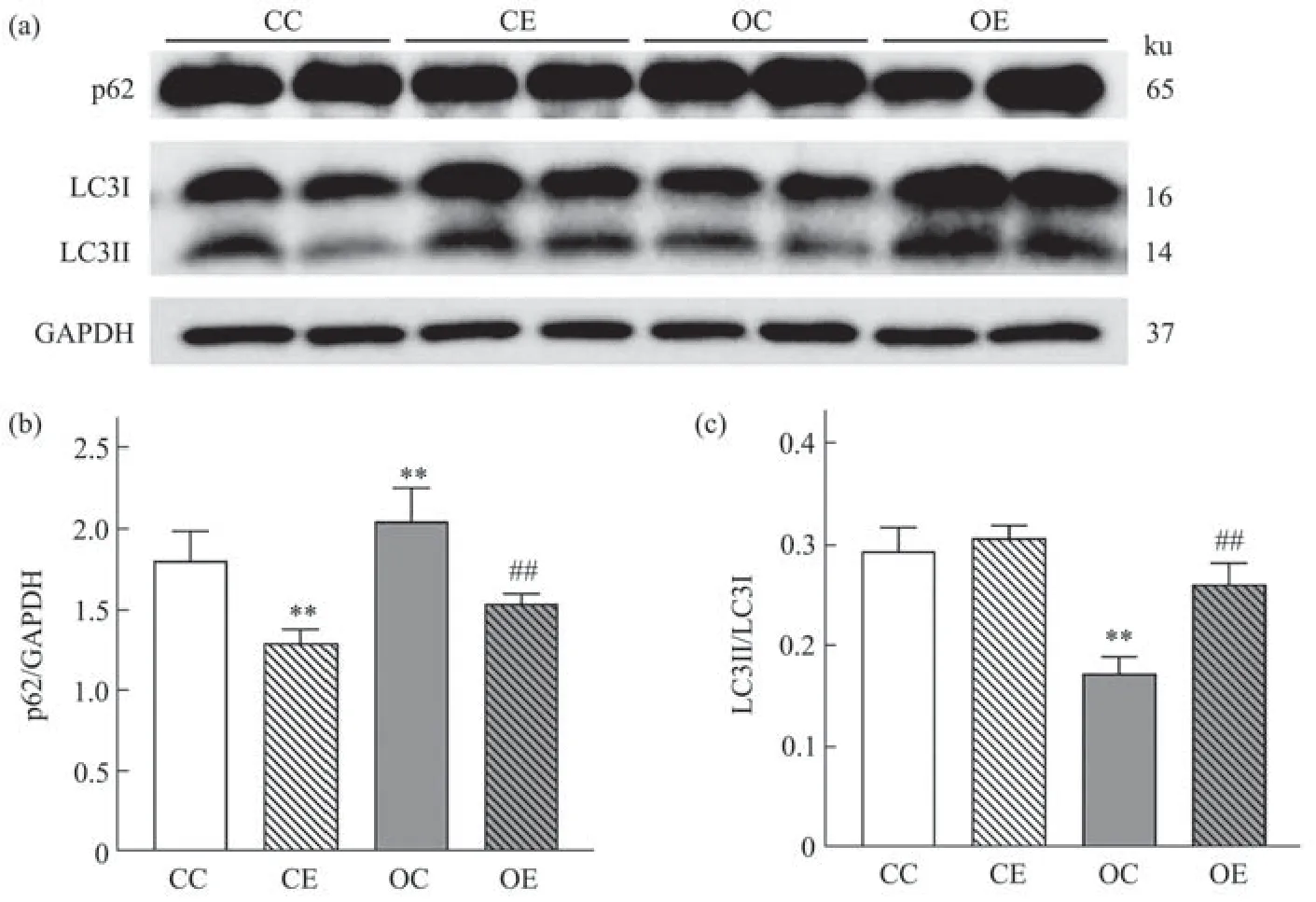
Fig. 5 Comparison of p62 and LC3II/LC3I in each group
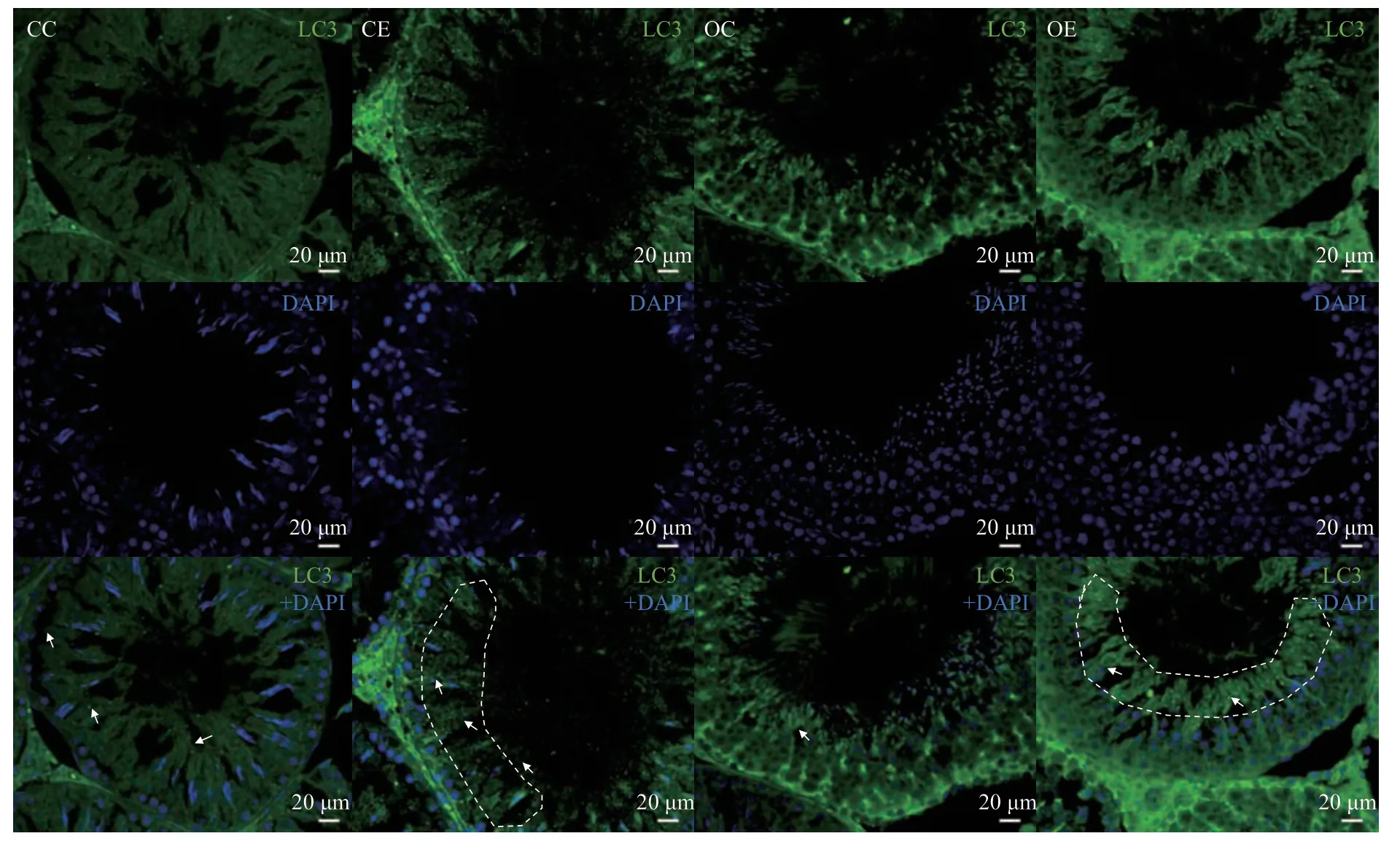
Fig. 6 Immunofluorescence of LC3 protein in testicular tissue in each group
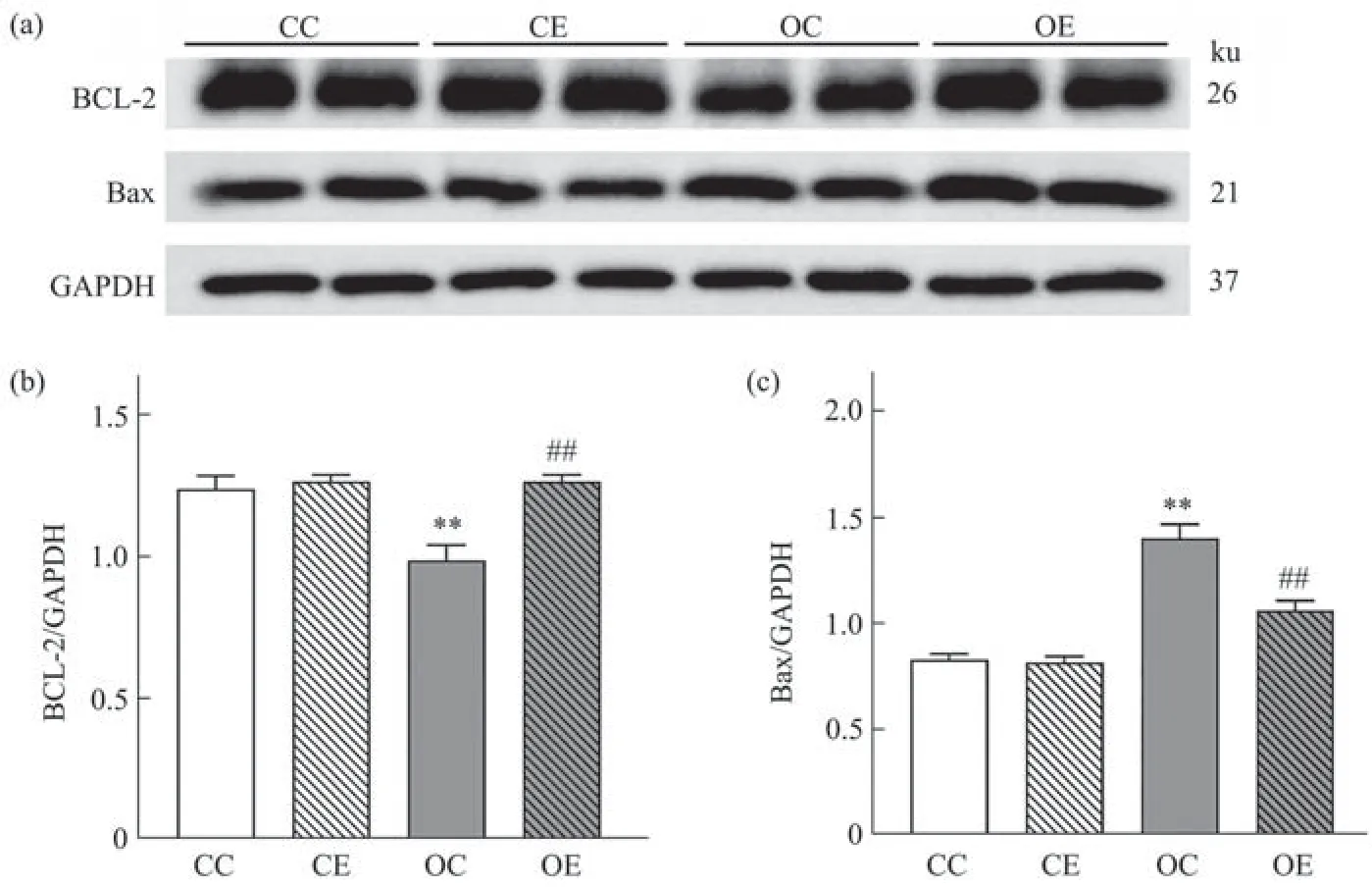
Fig. 7 Comparison of BCL-2 and Bax in each group

Fig. 8 Immunofluorescence of BCL-2 protein in testicular tissue in each group

Fig. 9 Comparison of AMPK in each group

Fig. 10 The correlation between AMPK and Bax, p62,BCL-2, and LC3 in the testicular tissue
2.2 运动干预对肥胖大鼠体重及睾丸指数的影响
8周干预后,与CC组相比较,OC组大鼠体重升高,CE组大鼠体重降低,与OC组相比较,OE组大鼠体重降低,均具有统计学显著意义(图2a,P<0.01)。双因素方差分析结果表明,高脂饮食(F(1,20)=40.395,偏η2=0.669,P<0.001)和运动(F(1,20)=44.225,偏η2=0.689,P<0.001)分别对大鼠体重影响的主效应效果显著,但二者对于体重的影响不存在交互作用(F(1,20)=1.314,偏η2=0.062,P=0.265)。与CC组相比较,CE组大鼠睾丸指数增加(P<0.05),OC组大鼠睾丸指数有降低趋势,但无显著性差异(P>0.05)(图2b)。双因素方差分析结果高脂饮食(F(1,20)=5.951,偏η2=0.229,P=0.024)和运动(F(1,20)=7.043,偏η2=0.26,P=0.015)对睾丸指数影响的主效应显著,但高脂饮食和运动对大鼠睾丸指数影响不存在交互作用(F(1,20)=0.589,偏η2=0.029,P=0.452)。这表明运动与饮食对体重与睾丸指数均不存在交互作用,主效应分析显示运动与高脂饮食分别对体重与睾丸指数的影响具有统计学意义。
2.3 运动干预对肥胖大鼠睾丸形态结构的影响
通过观察睾丸HE染色结果(图3)发现,与CC组相比,OC组睾丸组织各级生精细胞排列混乱,生精细胞层数和精原细胞数量降低,出现脂滴沉积,间质细胞和支持细胞数量有所减少,生精小管直径显著降低(图4a,P<0.05)。运动干预后,与OC组相比较OE组大鼠生精细胞层数有所增加,细胞间脂滴沉积减少,间质细胞和支持细胞数量有所增加,生精小管直径和精原细胞数量显著升高(图4,P<0.05)。双因素方差分析结果显示,高脂饮食(F(1,96)=0.073,偏η2=0.001,P<0.001)和运动(F(1,96)=158.92,偏η2=0.623,P<0.001)分别对于生精小管直径的主效应效果显著,但二者不存在交互作用(F(1,96)=2.501,偏η2=0.025,P=0.117)。对于精原细胞数量而言(图4b),高脂饮食的主效应效果不显著(F(1,96)=0.073,偏η2=0.001,P=0.787),运动的主效应显著(F(1,96)=158.92,偏η2=0.623,P<0.001),二者对大鼠精原细胞数量的影响不存在交互作用(F(1,96)=2.501,偏η2=0.025,P=0.117),表明运动干预能够显著升高肥胖大鼠生精小管直径和精原细胞数量。
2.4 运动干预对肥胖大鼠睾丸组织p62和LC3蛋白表达的影响
与CC组相比,OC组睾丸中p62表达水平升高,LC3II/LC3I降低,CE组大鼠睾丸中p62表达水平降低,与OC组相比,OE组大鼠睾丸中p62表达水平降低LC3II/LC3I升高,均具有统计学显著意义(图5,P<0.01)。双因素方差分析结果表明,高脂饮食对睾丸内p62(F(1,20)=54.36,偏η2=0.731,P<0.001)和LC3(F(1,20)=19.445,偏η2=0.493,P<0.001)蛋白表达的主效应均效果显著,运动干预对睾丸内p62(F(1,20)=208.968,偏η2=0.913,P<0.001)和LC3(F(1,20)=7.144,偏η2=0.263,P=0.015)蛋白表达的主效应均显著,高脂饮食和运动对睾丸内p62蛋白表达的影响具有交互作用(F(1,20)=26.407,偏η2=0.569,P<0.001),但对LC3蛋白表达的影响不存在交互作用(F(1,20)=3.891,偏η2=0.163,P=0.063)。提示高脂饮食和运动干预两个因素对自噬相关蛋白质均有显著影响,运动干预能够显著下调肥胖大鼠睾丸p62蛋白表达,提高LC3II/LC3I蛋白表达。
如图6所示,利用免疫荧光技术对睾丸组织中LC3蛋白进行定位后发现,LC3主要表达于生精小管内的精母细胞,精子细胞以及精子等生精细胞中,睾丸间质细胞中较少。与CC组相比较,OC组LC3蛋白数量减少;与OC组相比较,OE组大鼠睾丸LC3蛋白表达数量增加,且在精子位置表现出明显增加。说明运动干预主要提高肥胖大鼠精子细胞位置中的自噬活性。
2.5 运动干预对肥胖大鼠睾丸组织BCL-2和Bax蛋白表达的影响
与CC组相比较,OC组大鼠睾丸中BCL-2蛋白表达水平降低,Bax蛋白表达水平升高,与OC组相比较,OE组大鼠睾丸组织中BCL-2蛋白表达水平升高,Bax蛋白表达水平降低,均具有统计学显著意义(图7a~c,P<0.01)。双因素方差分析结果表明,高脂饮食对BCL-2(F(1,20)=4.362,偏η2=0.179,P=0.049)和Bax(F(1,20)=84.486,偏η2=0.809,P=0.001)蛋白表达影响的主效应显著,运动干预对BCL-2(F(1,20)=15.913,偏η2=0.443,P=0.001)和Bax(F(1,20)=15.873,偏η2=0.442,P=0.001)蛋白表达影响的主效应显著,且二者对BCL-2(F(1,20)=11.627,偏η2=0.368,P=0.003)和Bax(F(1,20)=13.512,偏η2=0.442,P=0.001)蛋白表达的影响均存在交互作用。提示高脂饮食和运动干预对于BCL-2和Bax蛋白均有显著影响,运动干预能够显著升高肥胖大鼠睾丸BCL-2蛋白表达,下调Bax蛋白表达。
如图8所示,利用免疫荧光技术对睾丸组织中BCL-2蛋白进行定位后发现,BCL-2蛋白主要表达于生精小管中生精细胞中。与CC组相比较,高脂饮食干预后OC组生精小管内BCL-2蛋白表达数量降低;与OC组相比较,OE组大鼠睾丸生精小管中BCL-2蛋白表达数量增加,尤其是在精母细胞和精子细胞中,增加效果明显。说明运动干预抑制肥胖大鼠睾丸精母细胞和精子细胞中的凋亡过程。
2.6 运动干预对肥胖大鼠睾丸组织AMPK蛋白表达的影响
与CC组相比较,OC组大鼠睾丸组织中AMPK表达水平显著降低,与OC组相比较,OE组AMPK蛋白表达水平显著升高,均具有统计学显著意义(图9a,b,P<0.01)。双因素方差分析结果表明,高脂饮食(F(1,20)=237.192,偏η2=0.922,P<0.001)和运动(F(1,20)=95.644,偏η2=0.827,P<0.001)干预对睾丸内AMPK蛋白表达的主效应效果均显著,且二者对睾丸内AMPK蛋白表达的影响具有交互作用(F(1,20)=0.456,偏η2=0.182,P=0.048)。
2.7 睾丸组织AMPK与自噬和凋亡相关蛋白变化关系
由大鼠睾丸组织中AMPK与自噬和凋亡相关蛋白质的相关关系图(图10)可知,饼状图中颜色面积越大,相关度越高。图中以颜色表示相关性,红色为负相关,蓝色为正相关。通过对所有大鼠睾丸组织中AMPK与p62、LC3II/LC3I、BCL-2和Bax蛋白表达的相关性分析,发现AMPK与p62、LC3II/LC3I、Bax蛋白表达均为强相关(P<0.001)。
3 讨论
高脂饮食和静坐少动的生活行为是导致肥胖发生的主要因素,过度的脂肪堆积导致机体出现能量失衡,在诱发代谢相关疾病的同时导致生殖功能障碍。运动作为促进脂肪分解的最佳手段之一,对肥胖哺乳动物生殖功能的改善也有积极的作用,而这一过程可能通过细胞自噬和凋亡介导[30-31],但机制尚不明确。本研究通过对肥胖大鼠进行8周中等强度运动干预,通过观察生精小管结构,并检测运动激活自噬与凋亡过程相关的蛋白质,探究运动对肥胖大鼠睾丸组织的影响及其可能机制。
肥胖或高脂饮食环境状态下,脂肪堆积会导致生殖器官、肝脏、骨骼肌等非脂肪组织的损伤从而导致其功能障碍[32-34]。在本实验中,高脂饮食诱导的肥胖大鼠在体重显著增加的同时,睾丸指数降低,睾丸组织中出现脂滴沉积,生精小管中精子数量明显减少。自噬虽有清除损伤细胞和消除脂滴的作用,但在营养过剩等正能量平衡条件下随AMPK的抑制、mTORC1的激活而受到抑制,导致自噬过程无法启动。本研究结果表明,肥胖大鼠睾丸组织在出现脂滴沉积、精子数量减少等现象的同时,LC3II/LC3I比值显著降低,p62蛋白水平明显升高,同时AMPK蛋白表达受到抑制。在自噬发生过程中,LC3I需要不断脂化形成LC3II,促进自噬体膜的形成,因此一般以LC3I向LC3II转化的比例作为判断自噬活性的标志[35]。张欣等[36]对大鼠进行高脂高糖饮食干预后也发现,附睾组织中自噬相关蛋白LC3表达水平降低,p62蛋白表达水平增高,自噬过程受到抑制。此外,本研究利用免疫荧光手段对睾丸组织中LC3蛋白表达位置进行检测发现,肥胖大鼠睾丸组织中LC3蛋白的减少主要发生在生精细胞中,尤其是在生精细胞和精子中出现明显的减少。提示高脂饮食带来的肥胖通过抑制生精细胞中的自噬活性来影响睾丸的功能。
激活自噬能够改善能量过剩条件下的睾丸组织损伤。冯劲宜等[37]发现经腹腔注射脂联素治疗的糖尿病小鼠睾丸p62 mRNA表达水平降低,LC3 mRNA表达水平升高,且生精小管形态有所改善,生精细胞及支持细胞数目明显增加。Gamberi等[38]通过细胞实验验证了脂联素可通过激活AMPK诱导骨骼肌细胞的自噬、抑制凋亡,促进肌细胞存活。本研究对肥胖大鼠进行8周中等强度运动干预后,大鼠体重显著降低、睾丸指数升高的同时,睾丸组织中脂滴沉积现象明显减少,生精细胞层数和精子数量增加,大鼠睾丸组织内AMPK和LC3II/LC3I比值升高,p62蛋白表达降低。夏俊等[39]对肥胖大鼠进行10周游泳运动干预后发现,游泳运动使大鼠睾丸组织内自噬标志物LC3II表达增多,p62累积减少,与本研究结果相一致。本研究通过免疫荧光定位还观察到,运动对于睾丸LC3蛋白影响作用显著,在运动干预后正常饮食和高脂饮食的大鼠睾丸组织中LC3蛋白含量在精子和精原细胞位置均表现出明显增加,同时HE染色结果也显示在运动刺激下生精小管中精原细胞数量会出现增加,提示运动后精原细胞的变化可能是由于自噬过程介导。而AMPK可能是运动激活自噬的关键因子,在本研究中,睾丸组织中AMPK的表达水平受运动干预影响显著,在运动刺激下正常饮食和高脂饮食诱导的肥胖大鼠睾丸组织中AMPK蛋白表达水平均出现明显升高,且与自噬相关蛋白均呈强相关。谢凌坚等[40]报道了6周高强度间歇运动能显著增加野生型小鼠骨骼肌细胞中AMPKα2的磷酸化水平,LC3II/LCI比值显著增加,p62表达显著减少,但在AMPKα2基因敲除的小鼠中自噬活性未明显增加,说明AMPK在运动激活自噬的过程中发挥重要作用,而在睾丸组织中运动也可能通过AMPK诱导睾丸组织自噬的发生。
自噬与细胞凋亡存在交互作用,肥胖在抑制睾丸组织自噬的同时也会诱导生殖细胞凋亡。在超重及肥胖男性不育患者中,精子早期凋亡率明显增高且身体质量指数(BMI)与精子活动率呈负相关性[41]。Zhu等[42]发现肥胖男性中BCL-2/Bax比值在基因与蛋白质表达均出现下调。在动物实验也发现,8周高脂饮食后大鼠睾丸生精小管直径、生精细胞及间质细胞计数降低的同时,BCL-2表达减少,Bax表达升高[43]。在本研究中也观察到,肥胖导致大鼠生精小管直径减少,各级生精细胞排列混乱的同时,睾丸组织内Bax表达增加,BCL-2表达减少,从免疫荧光检测结果来看,凋亡的发生位置主要在生精细胞中。说明肥胖通过激活凋亡信号通路促进生精细胞中凋亡相关因子的表达,导致生精障碍,从而影响生殖功能。
运动在改善肥胖哺乳动物细胞凋亡中发挥积极作用早有报道。前期研究发现,运动能够改善由于肥胖导致的骨骼肌和海马体中的细胞凋亡[44-45]。对肥胖小鼠进行8周的中等强度跑台运动可使睾丸曲细精管结构得到改善,Bax表达下调,BCL-2表达上升[46]。在本研究中,运动干预使肥胖大鼠睾丸组织内BCL-2蛋白表达上调,Bax蛋白表达下降,说明运动可抑制由肥胖导致的睾丸细胞凋亡现象。同时,本研究还发现,运动干预对于睾丸细胞的凋亡的抑制作用主要针对肥胖大鼠,运动干预后正常饮食大鼠睾丸中凋亡相关蛋白质表达水平未发生显著变化,而肥胖大鼠睾丸组织中的凋亡现象则得到明显抑制,因此猜测运动干预主要在肥胖或高脂饮食环境下对于生殖器官的凋亡起到矫正作用。在睾丸组织中,AMPK除参与启动自噬过程外,也可能参与运动调控凋亡的过程,在本研究中AMPK与Bax呈强相关。Mu等[47]对肥胖诱导的精子生成障碍的小鼠模型注射重鸢尾素后,发现鸢尾素可激活AMPKα信号通路以减少睾丸细胞的凋亡,当特异性敲除AMPKα后,鸢尾素抑制睾丸凋亡的作用消除。推测运动刺激后AMPK可能在睾丸细胞自噬和凋亡过程中同时发挥作用,但其机制有待进一步探究。
4 结论
高脂饮食诱导的肥胖可导致大鼠睾丸组织形态结构的改变,并伴随自噬活性降低,凋亡增加。而8周中等强度运动干预可有效提高肥胖大鼠睾丸组织自噬水平并抑制细胞凋亡,改善生精小管结构,且这一过程可能通过运动激活AMPK介导。