桃小食心虫成虫期高表达气味结合蛋白基因的克隆与表达谱分析
2022-12-28李凯旋李艳艳王树娟张丽莹庞保平
李凯旋, 李艳艳, 王树娟, 斯 琴, 张丽莹, 庞保平
(1. 内蒙古农业大学草原昆虫研究中心, 呼和浩特 010020; 2. 呼和浩特市园林植保站, 呼和浩特 010030; 3. 呼和浩特市成吉思汗公园, 呼和浩特 010010)
桃小食心虫Carposinasasakii属鳞翅目(Lepidoptera)蛀果蛾科(Carposinidae),在我国分布范围广,主要危害苹果、枣、梨、桃、李、杏、山楂等多种果树的果实,每年造成果树落果和虫果率高达50%,是果园中危害最大、发生最普遍的主要害虫之一(孙丽娜等, 2015)。桃小食心虫初孵幼虫蛀果后,整个幼虫时期均在果内为害,直至老熟幼虫脱果后,落入土中结茧化蛹。目前我国对桃小食心虫的防控方法,主要以套袋和化学防治为主,由于用药量多、用药品种泛滥、种植模式调整、全球气候变暖等原因,桃小食心虫的危害大幅回升,抗药性发展加快。因此,桃小食心虫可能再次成为严重危害果树的重要害虫。
昆虫对外界气味分子的识别是一个非常复杂的过程,目前普遍的说法是气味分子通过昆虫嗅觉感器表面的小孔进入触角感器,与气味结合蛋白(odorant-binding protein, OBP)特异结合,穿过嗅觉感器淋巴液,最终激活结合受体,受体被气味分子激活后,化学信号转变为电信号,引发一系列神经活动,指导昆虫产生各种行为反应(Rützler and Zwiebel, 2005; 杨安等, 2019; 彭竹等, 2021)。参加昆虫嗅觉识别过程的气味蛋白主要包括气味结合蛋白(OBP)、化学感受蛋白(chemosensory protein, CSP)、气味降解酶(odorant-degrading enzyme, ODE)、气味受体(odorant receptor, OR)、离子型受体(ionotropic receptor, IR)和感觉神经元膜蛋白(sensory neuron membrane protein, SNMP)(Leal, 2013)。其中,OBPs是受体ORs与外界环境的重要载体,OBPs对化学物质的识别被认为是气味分子对昆虫行为产生影响的第一步。OBPs作为昆虫体内最早被发现并开始研究的嗅觉结合蛋白,已经在双翅目、鳞翅目、鞘翅目等昆虫中被发现,到目前被鉴定的OBPs超过600多个(Pelosietal., 2006, 2018; Zhouetal., 2010)。OBPs是一种小分子酸性、水溶性蛋白,主要存在于触角淋巴液中(Pelosi and Maida, 1995),分子质量为15~17 kD,大约由120~160个氨基酸组成。根据结合的气味分子不同,OBPs可以分为性信息素结合蛋白(pheromone-binding protein, PBP)、普通气味结合蛋白(general odorant-binding protein, GOBP)和触角结合蛋白(antennal-binding protein x, ABPx)(王桂荣等, 2002)。目前,OBPs功能已经通过分子生物学、生物化学、生物物理学、结构生物学、动力学和电生理学等手段逐步解析(Sandleretal., 2000; Zhangetal., 2017; Zhuetal., 2017)。通过qRT-PCR技术,研究发现OBPs不仅在触角中表达,同时在其他组织中也有表达,例如在头、下唇须、喙、腹、足、性腺中等,此外在幼虫及蛹期也有表达,根据基因表达与功能的联系,研究认为在触角中特异性或高丰度表达的OBPs主要参与昆虫的嗅觉识别(De Biasioetal., 2015; Yuanetal., 2015)。免疫定位和原位杂交等技术进一步明确了每种感器一般只表达单一类型的OBPs,例如,PBPs在毛形感器中专一表达,GOBPs则大多数在锥形感器中表达(Vogtetal., 2002)。
昆虫嗅觉识别机制的解析,对害虫生物防治具有重要的意义。对昆虫嗅觉识别机制的探讨, 有利于寻找新的化合物,通过干扰昆虫嗅觉识别,进而达到对靶标害虫的有效控制。桃小食心虫成虫期是控制此害虫发生为害的关键时期。因此,解析桃小食心虫成虫嗅觉识别的分子机制,对于制定有效的绿色防控措施具有重要的意义。基于此,本研究根据前期获得的转录组数据(Tianetal., 2018),筛选在桃小食心虫雌雄成虫触角中相对表达量较高的5个气味结合蛋白(OBP)基因(CsasOBP7,CsasOBP12,CsasOBP15,CsasOBP19和CsasOBP21),采用RACE克隆技术对其cDNA全长序列进行克隆,并对这5个CsasOBP基因在桃小食心虫不同发育阶段及刚羽化、交配高峰期及交配后6 h成虫不同组织内的表达量进行测定,以明确桃小食心虫发育及交配过程中关键气味结合蛋白的表达模式,为制定利用基因编辑等方式定向调控桃小食心虫的嗅觉行为的新型防治策略提供理论依据。
1 材料与方法
1.1 供试材料
供试桃小食心虫采自中国农科院果树研究所(40.61°N, 120.73°E),实验室置于温度(25±1)℃、相对湿度(70±5)%、光周期15L∶9D的人工气候箱中饲养。昆虫饲养方法:将卵卡放置于未成熟金冠苹果上,初孵幼虫蛀果后,将虫果置于上述培养箱中,待老熟幼虫脱果后放入木屑中化蛹,羽化后成虫饲喂10%蜂蜜水。
1.2 RNA提取及cDNA合成
取100对桃小食心虫成虫触角,提取RNA用于CsasOBP基因全长克隆。为明确CsasOBP基因在桃小食心虫不同发育阶段及成虫交配前后的表达情况,实验选取完整的桃小食心虫1日龄卵、5日龄卵、初孵幼虫、老熟幼虫和蛹(3日龄)迅速置于液氮中。分别切取刚羽化、交配高峰期和交配后6 h桃小食心虫雌雄成虫触角、头(不含触角)、胸、腹、足及翅样品,每个样品取50~60头。按照MiniBEST Universal RNA Extraction Kit (TaKaRa,大连),总RNA提取试剂盒说明书提取上述所有样品总RNA,用超微量紫外分光光度计(IMPLEN-P330,德国)和1%琼脂糖凝胶电泳检测总RNA的浓度及质量。根据反转录试剂盒说明书合成cDNA第1链,在-20℃冰箱保存备用。
1.3 RACE全长基因克隆
基于前期桃小食心虫转录组测序数据(Tianetal., 2018),筛选并获得在桃小食心虫雌雄成虫触角中相对表达量较高的5个CsasOBP基因(CsasOBP7,CsasOBP12,CsasOBP15,CsasOBP19和CsasOBP21)的片段序列,设计目的基因序列3′RACE和5′RACE片段特异性引物(表1)。中间片段扩增PCR反应体系: 成虫触角cDNA模板1 μL, 正反向引物(10μmol/L)各1 μL, Premix TaqTM(V2.0 plus Dye)(TaKaRa,大连)12.5 μL, ddH2O 9.5 μL。反应程序: 94℃预变性3 min; 94℃变性30 s, 50℃ 退火30 s, 72℃ 延伸1 min, 30个循环;72℃延伸10 min。1.5%琼脂糖凝胶电泳检测扩增产物,用 DNA凝胶回收试剂盒回收纯化目的片段,连接pMD19-T克隆载体,转化到大肠杆菌Escherichiacoli感受态细胞DH5α后涂布含有氨苄的LB固体培养基上,37℃培养12 h,挑取白色单菌落进行PCR验证后再将验证正确的菌液于LB液体培养基中过夜培养,送样测序。
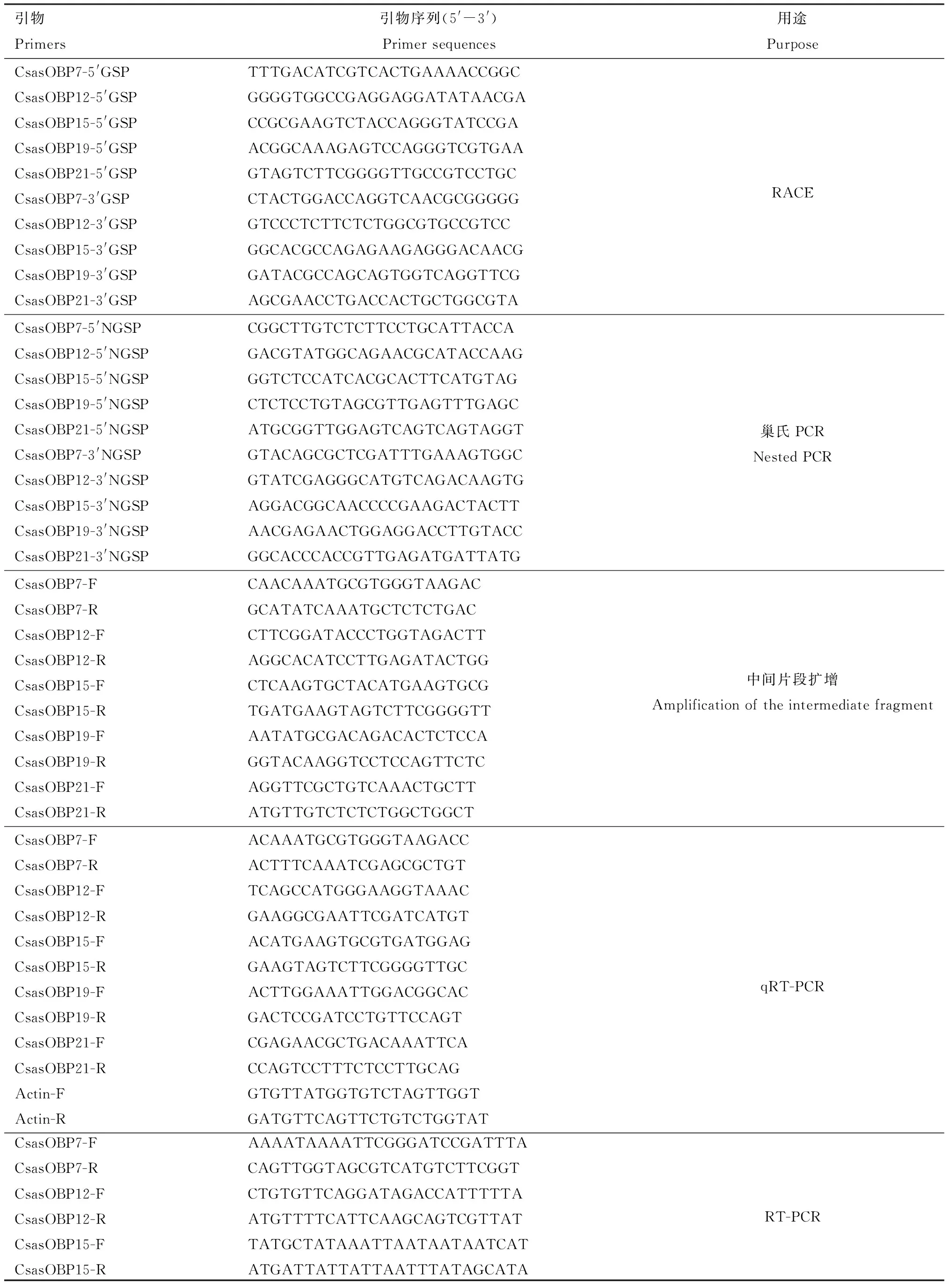
表1 引物信息Table 1 Primer information

续表1 Table 1 continued
CsasOBP基因的3′末端和5′末端序列扩增:根据测序验证结果设计3′-和5′-特异性引物(表1),按照SMARTer®RACE 5′/3′ Kit试剂盒(TaKaRa,大连)说明书进行扩增,巢氏PCR程序: 94℃ 30 s, 72℃ 2 min, 5个循环; 94℃ 30 s, 70℃ 30 s, 72℃ 2 min, 5个循环; 94℃ 30 s, 68℃ 30 s, 72℃ 2 min, 20个循环。再以扩增产物为模板进行巢氏PCR: 94℃ 30 s, 68℃ 30 s, 72℃ 2 min, 25个循环。得到扩增PCR产物处理方法同上。测序得到的3′与5′端序列片段与中间片段拼接,最终得到全长cDNA序列。
1.4 基因序列生物信息学分析
利用DNAMAN 6.0软件将翻译后的基因序列与其他昆虫的OBP基因序列进行比对;使用在线程序Expasy(http:www.expasy.org/tools/pi_tool.html)对5个CsasOBPs的分子量及理论等电点进行预测;使用Signal P 5.0(https:∥services.healthtech.dtu/service.php?SignalP-5.0)预测5个CsasOBPs的信号肽序列。运用NCBI Conserved Domains(http:∥www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)分析5个CsasOBPs的保守结构域;用ProtScale(http:∥web.expasy.org/ cgi-bin/protscale/protscale.pl)对5个CsasOBPs进行疏水性分析;使用PredictProtein(https:∥www.predictprotein.org/)预测氨基酸序列中的二硫键位置和蛋白二级结构;运用在线软件Swiss-Model(https:∥www. swissmodel.expasy.org/)对蛋白三级结构进行预测;在NCBI中利用Blast进行5个CsasOBP蛋白的同源搜索和比对(e值设定为10-5),选择其他鳞翅目昆虫的气味结合蛋白氨基酸序列,经Clustal X进行多序列比对。 使用MEGA 6.0软件的邻接法(neighbor-joining method, NJ method)(重复运行1 000次)构建系统发育树。
1.5 基因时空表达测定
基于1.3节克隆的5个CsasOBP基因序列,利用Primer Premier 5.0设计半定量RT-PCR引物(表1), 利用PrimerBank在线软件(https:∥pga.mgh.harvard.edu/primerbank/)设计qRT-PCR引物(表1),选用Actin作为内参基因,以1.2节合成的cDNA为模板,每个样品3次生物学重复,4次技术重复。半定量RT-PCR反应体系(25 μL): Premix TaqTM(V2.0 plus Dye) 12.5 μL, cDNA模板1 μL, 正反向引物(10 μmol/L)各1 μL, Nuclease-Free Water 9.5 μL。RT-PCR反应程序: 94℃预变性3 min; 94℃变性 30 s, 55℃退火 30 s, 72℃延伸2 min, 35次循环;72℃延伸10 min。1.5%琼脂糖凝胶电泳检测表达量。qRT-PCR反应体系(20 μL): SYBRⅡMix 10 μL, cDNA模板2 μL,正反向引物(10 μmol/L)各0.4 μL, Nuclease-Free Water 7.2 μL。qPCR反应程序: 95℃预变性10 min; 95℃变性15 s, 60℃退火15 s, 72℃延伸30 s, 40次循环。所有处理均以CsasOBP7在刚羽化的雌成虫触角中的表达量为对照,采用2-ΔΔCt法计算5个CsasOBP基因在不同发育阶段及不同组织的相对表达量。
1.6 数据分析
使用SPSS 20统计分析软件对数据进行分析,采用单因素方差分析和Duncan氏多重极差检验,对每个基因在不同发育阶段及在相同性别不同组织中的表达量进行差异显著性分析,利用独立样本T检验对每个基因在相同组织不同性别的表达量进行差异显著性分析。
2 结果
2.1 桃小食心虫OBP基因的序列特征
克隆获得桃小食心虫5个气味结合蛋白基因CsasOBP7,CsasOBP12,CsasOBP15,CsasOBP19和CsasOBP21全长cDNA序列(GenBank登录号分别为MZ476786-MZ476790)。这5个基因具有完整的开放阅读框,开放阅读框长度分别为432, 543, 411, 765和483 bp,分别编码143, 180, 136, 254和160个氨基酸,蛋白分子量分别为16.35, 19.94, 14.86, 29.01和18.48 kD,等电点分别为5.06, 6.88, 4.32, 6.91和5.11,均含有信号肽。序列比对(图1)发现,CsasOBP7, CsasOBP12, CsasOBP15和CsasOBP21均含有6个保守的半胱氨酸位点,第2-3个半胱氨酸残基之间间隔3个其他氨基酸残基,属于完整的Classical OBPs;CsasOBP19经对比分析后确定为C端不完整的Minus-C OBP。
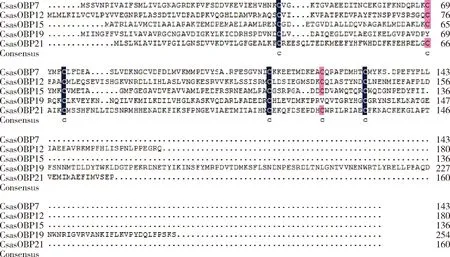
图1 桃小食心虫5个CsasOBPs的氨基酸序列比对Fig. 1 Amino acid sequence alignment of five CsasOBPs of Carposina sasakiiCsasOBPs的GenBank登录号GenBank accession numbers of CsasOBPs: CsasOBP7: MZ476786.1; CsasOBP12: MZ476787.1; CsasOBP15: MZ476788.1; CsasOBP19: MZ476789.1; CsasOBP21: MZ476790.1. 6个保守的半胱氨酸位点中,蓝色区域相似度为100%,粉色区域相似度≥75%。Among the six conservative cysteine sites, the similarity of blue area is 100% and the similarity of pink area is ≥75%.
通过DNAMAN比对发现CsasOBP7与大螟Sesamiainferens的SinfOBP16(GenBank登录号: AGS36756.1)的氨基酸序列一致性最高,为51.75%;CsasOBP12与马尾松毛虫Dendrolimuspunctatus的DpunOBP39(GenBank登录号: ARO70198.1)氨基酸序列一致性最高,为76.5%;CsasOBP15与沙棘木蠹蛾Eogystiahippophaecolus的EhipOBP (GenBank登录号: AOG12857.1)氨基酸序列一致性最高,为58.57%;CsasOBP19与二化螟Chilosuppressalis的CsupOBP(GenBank登录号: AGM38607.1)氨基酸序列一致性最高,为65.75%;CsasOBP21与小菜蛾Plutellaxylostella的PxylOBP35(GenBank登录号: AMR99729.1)氨基酸序列一致性最高,为58.48%。
2.2 桃小食心虫OBP基因的系统发育
采用邻接法对桃小食心虫5个CsasOBPs与鳞翅目其他12个科昆虫的OBPs构建系统发育树(图2),从图中可以看出9种螟蛾科昆虫、5种夜蛾科昆虫、3种尺蛾科昆虫、2种枯叶蛾科昆虫、2种草螟科昆虫以及卷叶蛾科、菜蛾科、斑蛾科、蛱蝶科、木蠹蛾科、蛀果蛾科、卷蛾科各1种昆虫被分为4大分支。这5个CsasOBPs与其他科的昆虫同源性表现不同,但与DNAMAN比对结果相同,如CsasOBP19与二化螟的CsupOBP聚为一个分支且自检值高达100,说明它们有保守相似的功能。CsasOBP21和CsasOBP15与其他昆虫的OBPs同源性比较低,自检值分别为25和12,且CsasOBP15单独聚为一个小分支,说明CsasOBP15与其他几个OBPs相比可能有不同的功能。这5个CsasOBPs之间分别聚类在3个分支中,其中CsasOBP7和CsasOBP19聚在一个分支,其他3个CsasOBPs分别聚在2个分支中,表明CsasOBP7和CsasOBP19亲缘关系较近且可能有相似的功能,其他3个CsasOBPs互相之间功能可能存在差异。

图2 邻接法构建的基于氨基酸序列的桃小食心虫CsasOBPs与其他鳞翅目昆虫OBPs的系统进化树(1 000次重复)Fig. 2 Phylogenetic tree of CsasOBPs of Carposina sasakii and OBPs from other lepidopteran insects constructed by the neighbor-joining method based on amino acid sequence (1 000 replicates)蛋白来源物种Origin species of proteins: Pxyl: 小菜蛾Plutella xylostella; Sexi: 甜菜夜蛾Spodoptera exigua; Hrho: 重阳木斑蛾Histia rhodope; Cpun: 桃蛀螟Conogethes punctiferalis; Obru: 冬尺蠖蛾Operophtera brumata; Dple: 黑脉金斑蝶Danaus plexippus plexippus; Dpun: 马尾松毛Dendrolimuspunctatus; Csup: 二化螟Chilo suppressalis; Sinf: 大螟Sesamia inferens; Ehip: 沙棘木蠹蛾Eogystia hippophaecolus; Sins: 小线角木蠹蛾Streltzoviella insularis; Adis: 双委夜蛾Athetis dissimilis; Dabi: 冷杉梢斑螟Dioryctria abietella; Acon: 苹果果蛾Argyresthia conjugella; Msep: 玉米黏虫Mythimna separate; Dkik: 思茅松毛虫Dendrolimus kikuchii; Dhou: 云南松毛虫Dendrolimus houi; Eobl: 茶尺蠖Ectropis obliqua; Gpyl: 桑螟Glyphodes pyloalis; Hass: 烟草夜蛾Helicoverpa assulta; Cpin: 松蛀螟Conogethes pinicolalis; Gmel: 大蜡螟Galleria mellonella; Cmed: 稻纵卷叶螟Cnaphalocrocis medinalis; Gmol: 梨小食心虫Grapholita molesta; Scin: 国槐尺蠖Semiothisa cinerearia; Lbot: 葡萄花翅小卷蛾Lobesia botrana; Gcae: 黄翅绢丝野螟Glyphodes caesalis.
2.3 桃小食心虫OBP基因的时空表达
qRT-PCR结果表明,桃小食心虫的5个CsasOBP基因在不同发育阶段的表达量不同。CsasOBP7和CsasOBP19均在蛹期表达量最高(P<0.05),其中CsasOBP7在蛹期的表达量显著高于其他发育阶段的(P<0.05),而CsasOBP19除了在蛹期高表达外,在卵期表达量也较高,仅次于蛹期的。CsasOBP12,CsasOBP15和CsasOBP21均在卵期表达量最高(P<0.05),其中CsasOBP21除了在卵期高表达外,在蛹期的表达量也较高,仅次于卵期的,而CsasOBP15除了在卵期高表达外,在其他发育阶段表达量均无显著差异(P>0.05)(图3)。上述结果与半定量RT-PCR结果(图4)一致。
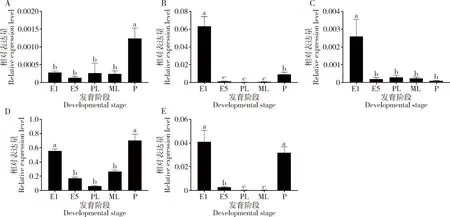
图3 桃小食心虫不同发育阶段5个CsasOBP基因表达量的qRT-PCR分析Fig. 3 Expression levels of five CsasOBP genes in Carposina sasakii at different developmental stages detected by qRT-PCRA: CsasOBP7: B: CsasOBP12; C: CsasOBP15; D: CsasOBP19; E: CsasOBP21. E1: 1日龄卵1-day-old egg; E5: 5日龄卵5-day-old egg; PL: 初孵幼虫Newly hatched larva; ML: 老熟幼虫Mature larva; P: 蛹Pupa. 图中数据为平均值±标准误;柱上不同字母表示差异显著(P<0.05, Duncan氏多重极差检验)。Data in the figure are mean±SE. Different letters above bars indicate significant difference (P<0.05, Duncan’s multiple range test).
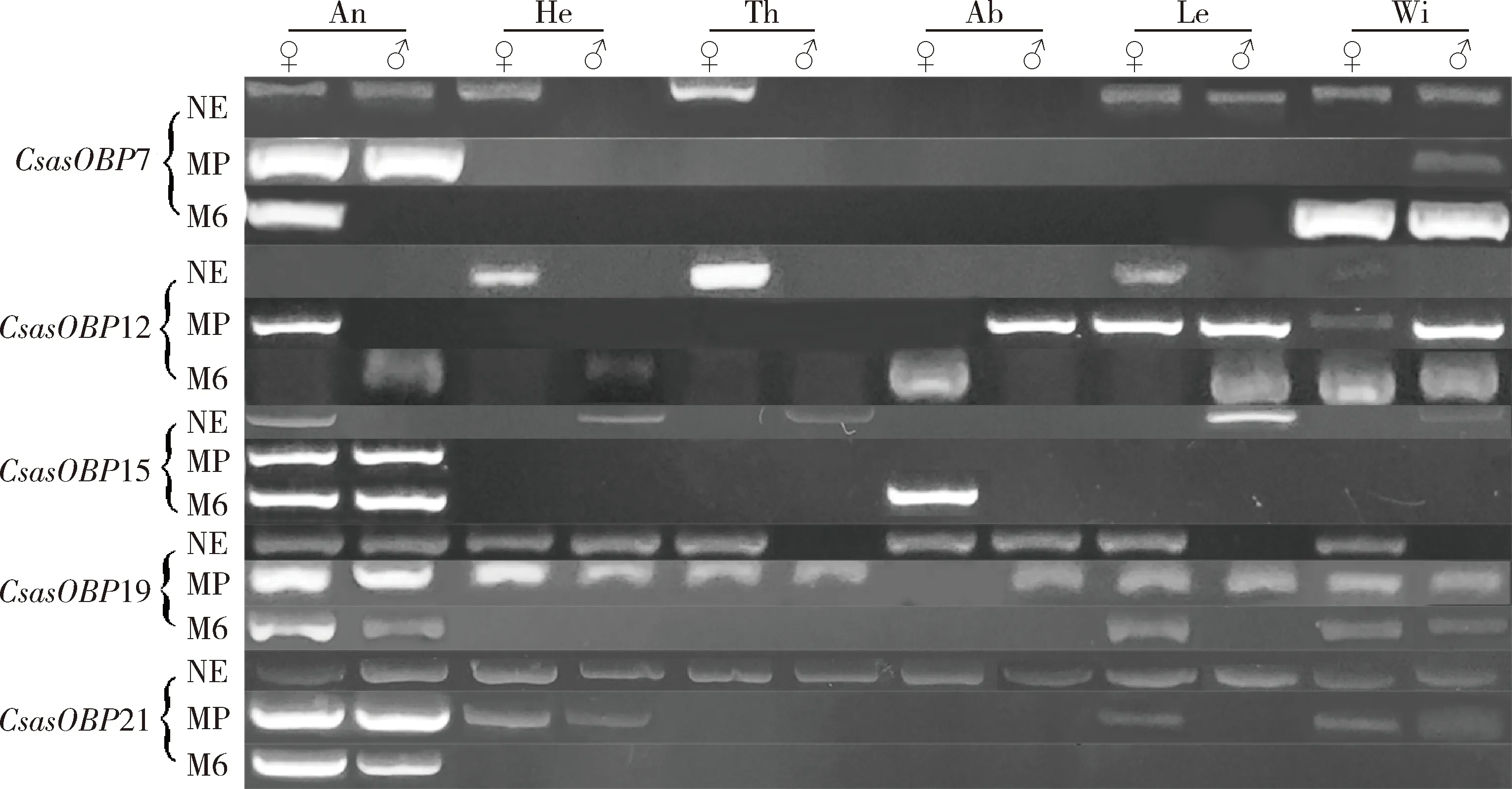
图4 桃小食心虫不同发育阶段成虫不同组织中5个CsasOBP基因表达量的半定量RT-PCR分析Fig. 4 Semi-quantitative RT-PCR analysis of the expression levels of five CsasOBP genes in different tissues of Carposina sasakii adults at different developmental stagesAn: 触角Antenna; He: 去除触角的头Head with antennae removed; Th: 胸Thorax; Ab: 腹Abdomen; Le: 足Leg; Wi: 翅Wing. NE: 刚羽化成虫Newly emerged adult; MP: 交配高峰期成虫Adult at the mating peak; M6: 交配后6 h的成虫Adult at 6 h after mating.
在成虫刚羽化时,5个CsasOBP基因的表达量在不同组织间均存在显著差异,CsasOBP7,CsasOBP12和CsasOBP21的表达量在各组织中最高,其次为CsasOBP19的,最低为CsasOBP15的;除CsasOBP19在雌虫腹部的表达量显著高于在雄虫腹部的(P<0.01)外,其他4个CsasOBP基因的表达量均在雄虫组织中高于在雌虫组织中(图5)。在成虫交配高峰期,5个CsasOBP基因在雄虫触角中表达量均显著高于在其他组织中的(P<0.05);CsasOBP7和CsasOBP15只在触角中表达(P<0.05),且在雄虫触角中的表达量显著高于在雌虫触角中的(P<0.01);CsasOBP12在雄虫触角中表达量最高(P<0.05),其次为在腹部以及在雄虫足和翅中的,在其他组织中不表达;CsasOBP19在雄虫触角中表达量最高(P<0.05),其次为在雌虫触角、足、翅以及雄虫头部(不含触角)和翅中,在胸部和腹部无表达;CsasOBP21 在雄虫触角中的表达量显著高于其他CsasOBP的,大约是其他4个CsasOBP基因最高表达量的70~1 900 倍(图6)。在成虫交配后6 h,CsasOBP19在雄虫触角和雌虫翅中高表达,CsasOBP21在触角中高表达且在雄虫触角中表达量显著高于雌虫触角中的(P<0.01),两者在其他组织中不表达或微量表达;CsasOBP7在雄虫足和翅中高表达(P<0.05),在其他组织中不表达或微量表达;CsasOBP12在雄虫腹部高表达(P<0.05),其次为在雄虫翅中,在其他组织中不表达或微量表达;CsasOBP15在雌虫腹部高表达(P<0.05),其次为在雄虫触角中,在其他组织中不表达(图7)。
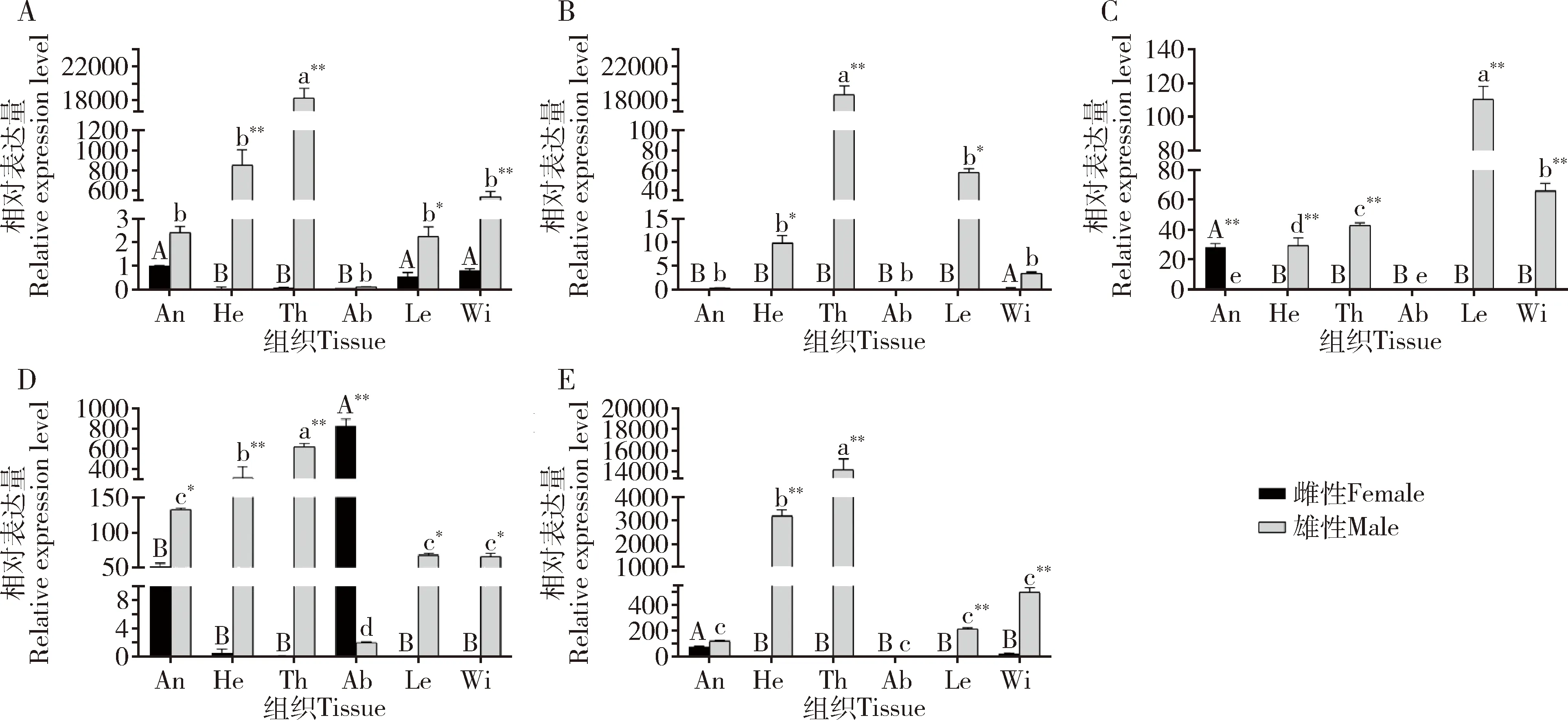
图5 5个CsasOBP基因在桃小食心虫成虫刚羽化时期不同组织中的表达量Fig. 5 Expressions levels of five CsasOBP genes in different tissues of the newly emerged adults of Carposina sasakiiA: CsasOBP7; B: CsasOBP12; C: CsasOBP15; D: CsasOBP19; E: CsasOBP21. An: 触角 Antenna; He: 去除触角的头 Head (excluding antenna); Th: 胸Thorax; Ab: 腹Abdomen; Le: 足Leg; Wi: 翅Wing. 图中数据为平均值±标准误;柱上星号表示同一组织雌雄间的基因表达量差异显著(*P<0.05; **P<0.01)(T检验); 柱上不同大写和小写字母分别表示基因表达量在雌和雄成虫不同组织间差异显著(P<0.05, Duncan氏多重极差检验)。图6和7同。Data in the figure are mean±SE. Asterisks above bars indicate significant difference in the gene expression level in the same tissue between female and male adults (*P<0.05; **P<0.01) (T-test). Different capital and lowercase letters above bars indicate significant difference in the gene expression level among different tissues of female and male adults, respectively (P<0.05, Duncan’s multiple range test). The same for Figs. 6 and 7.

图6 5个CsasOBP基因在桃小食心虫交配高峰期成虫不同组织中的表达量Fig. 6 Expressions levels of five CsasOBP genes in different tissues of Carposina sasakii adults at the mating peak stageA: CsasOBP7; B: CsasOBP12; C: CsasOBP15; D: CsasOBP19; E: CsasOBP21.

图7 5个CsasOBP基因在桃小食心虫交配后6 h成虫不同组织中的表达量Fig. 7 Expressions levels of five CsasOBP genes in different tissues of Carposina sasakii adults at 6 h after matingA: CsasOBP7; B: CsasOBP12; C: CsasOBP15; D: CsasOBP19; E: CsasOBP21.
3 讨论
本研究根据桃小食心虫转录组数据库,首次克隆了桃小食心虫5个CsasOBP基因cDNA全长序列。序列分析表明,CsasOBP7, CsasOBP12, CsasOBP15和CsasOBP21均含有6个保守的半胱氨酸,且第2-3个半胱氨酸之间有3个其他氨基酸,属于Classical OBPs;CsasOBP19为一段C端不完整的Minus-C OBP。与其他昆虫OBP序列同源对比和系统发育关系发现(图2),CsasOBP19和二化螟的CsupOBP聚类在同一分支;序列比对中也发现两个OBPs同源性最高。这也说明物种亲缘关系越近,气味蛋白间的序列相似性越高,同时也说明这两种气味结合蛋白在桃小食心虫和二化螟在嗅觉识别过程中可能有相似的功能。
绝大部分OBP基因在昆虫触角中特异性或高表达(陈秀琳等, 2018; 胡军等, 2019),这与OBPs作为载体参与触角淋巴液脂溶性气味分子运输的功能 (Bentonetal., 2007)相一致;同时OBP在雌雄虫的相同组织也有明显的偏向表达,可能与不同性别在昆虫特定时期的承担角色不同有关,如小菜蛾PxylOBP31在雄蛾触角中的表达量是雌蛾触角表达量的近2倍(覃江梅等, 2016)。在本研究中,虽然5个CsasOBP基因在桃小食心虫刚羽化时在成虫各组织中均有表达(图5),但在交配高峰期均在触角中特异性高表达(图6),且在雄虫触角中远高于在雌虫触角,这意味着这些OBP可能结合桃小食心虫雌成虫挥发的性信息素,在雄虫寻找雌虫过程中起着重要的作用。特别是CsasOBP21和CsasOBP7在雄虫触角中的表达量远高于其他3个CsasOBP基因,值得进一步研究。
本研究发现桃小食心虫刚羽化、交配高峰以及交配后6 h 3个时期(图5~7),5个CsasOBP基因的表达量总体上呈由高到低变化,这与桔小实蝇Bactroceradorsalis的BdorOBP61在桔小实蝇成虫各个时期表达情况 (龚妃良, 2014)一致。OBPs存在的组织与其功能有关(Lietal., 2008)。桃小食心虫雌雄成虫在求偶时均会产生振翅行为,同时雌虫求偶时前、中、后足会将整个虫体抬起与支持物平行(蒋天小, 2017)。CsasOBP12,CsasOBP19和CsasOBP21在交配高峰期的翅和足中表达较高可能与雌雄虫在交配高峰期的求偶行为有关。CsasOBP19在交配后6 h,在触角及翅中表达量较交配前升高且在雌成虫翅中大量表达,类似现象也出现在棉铃虫Helicoverpaarmigera中(李兆群等, 2017)。
综上所述,本研究成功克隆了桃小食心虫5个CsasOBP基因的全长cDNA序列,并对其生物信息学及表达特性进行了分析。 研究结果表明5个CsasOBP基因可能在桃小食心虫雄虫寻找雌虫过程中发挥着重要作用,为进一步研究其在桃小食心虫嗅觉识别中的功能,及其在桃小食心虫寻找配偶、寄主定位及产卵地选择等方面的作用奠定了必要的基础。
