林地植被类型变化对土壤真菌群落结构的影响
2022-09-26周红敏徐肇友骆先有陈杏林陈焕伟
周红敏,彭 辉,徐肇友,骆先有,陈杏林,陈焕伟
(1.龙泉市林业科学研究院,浙江 龙泉 323700;2.凤阳山管理处,浙江 龙泉 323700)
土壤真菌是土壤微生物的重要组分之一,在有机质分解,物质循环等方面发挥着重要作用[1-2]。真菌对土壤环境的变化较细菌更为灵敏[3-4],其群落结构变化被当作生态系统恢复的重要指标[5]。真菌群落结构变化跟土地利用方式[6]、管理措施[7-8]、pH 值[9]等有关。杉木Cunninghamia lanceolata是我国南方重要的经济树种[10-11]。相关研究表明上连栽会使土壤肥力下降[12],碳、氮循环菌减少等[13]。油茶Camellia oleifera是世界四大木本油料树种之一,是我国特有的食用木本油料作物[14-17]。在我国南方丘陵地区油茶林多是在杉木采伐迹地垦造完成,受油茶林管理及枯落物分解等影响,林地土壤环境等因子势必发生变化。目前,已有很多学者开展了,不同林龄油茶林化学计量特性[18-19]、土壤理化性质[20]等变化规律的研究,例如费裕翀[21]等研究了不同施肥处理对油茶林土壤微生物群落结构的影响。但对于杉木林改为油茶林后土壤真菌群落结构组成及多样性变化尚未见报道。因此,通过Illumina Miseq 测序技术,研究杉木林转变为油茶林后土壤真菌的变化情况,以期为油茶林的培育供科学依据。
1 研究区概况
研究区位于浙江省龙泉市上圩林区(27°59′05″~27°59′15″N,119°03′32″~19°04′17″E),属于中亚热带季风气候,年均降水量为1 650 mm 左右,年平均相对湿度79%。极端最高气温为41.5℃,极端最低气温为-8.5℃。油茶为2010年在杉木采伐迹地造林,胸径11.7 cm,树高3 m。油茶林每年抚育2 次,施复合肥1 125 kg/hm2。杉木林为1986年造林,胸径、树高分别为22.3 cm、17 m,近10年来杉木林除2019年施过复合肥750 kg/hm2以及深耕除草一次外,未进行任何人工管理。
2 研究方法
2.1 土壤样品采集
选取立地条件相近的杉木林和油茶林,于2020年10月在各林分中分别设置20 m×20 m 的3 块标准样地,采用5 点取样法采集0~20 cm 的表层土壤,将每一样地5 个取样点的土样混匀,挑出石砾、根系等杂质,其中一份装入无菌自封袋中于冷藏箱带回,置于-80℃冰箱中保存,用于土壤真菌群落结构多样性测定;另一份风干后过2 mm 筛用于土壤化学性质分析,土壤理化性质参照鲁如坤[22]的分析方法进行。
2.2 土壤DNA 提取和高通量测序
取0.5 g 土壤样品用于土壤总DNA 抽提(E.Z.N.A.® Soil DNA Kit 试剂盒),抽提后的DNA 利用 1% 琼脂糖凝胶电泳检测,DNA 的纯度和浓度采用NanoDrop2000进行检测;引物为SSU0817F(5′-TTAGCATGGA ATAATRRAATAGGA-3′) 和1196R(5′-TCTGGA CCTGGTGAGTTTCC-3′),扩增区域为V5-V7 区。用2%的琼脂糖凝胶对同一样本混合后PCR 产物进行电泳检测,PCR 产物回收采用AxyPrepDNA凝胶回收试剂盒,然后洗脱(Tris_HCl)、电泳检测(琼脂糖2%)。用Promega 公司的QuantiFluor ™-ST 蓝色荧光定量系统对PCR 产物进行检测定量。通过Illumina 公司的Miseq PE300平台进行测序(上海美吉生物医药科技有限公司)。
2.3 数据分析
所得数据用fastp(0.19.6)软件[23]进行数据质控,采用Mothur(1.30.2)软件进行Alpha 多样性分析(Simpson 指数、ACE 指数、Coverage 指数、Chao1 指数、Shannon 指数),Qiime(1.9.1)分析真菌各分类学分度,统计真菌在门、纲、属水平上的群落组成进行分析。以冗余分析(Redundancy analysis,RDA)分析土壤理化性质和优势真菌群落相关性。采用SPSS19.0 软件进行单因素分析和LSD 显著性检验,Excel 2016 软件进行数据统计和作图。
3 结果与分析
3.1 土壤理化性质
由表1可知,杉木林的土壤含水率、全氮、全磷、碱解氮和有机质的含量分别为24.63%、1.02 g/kg、0.26 g/kg、61.28 mg/kg 和38.19 g/kg,均显著高于油茶林(P<0.05)。杉木林土壤中全钾和pH 和油茶林相比差异不显著(P>0.05)。油茶林土壤有效磷和速效钾分别是杉木林的1.88倍和1.34 倍,显著高于杉木林(P<0.05)。
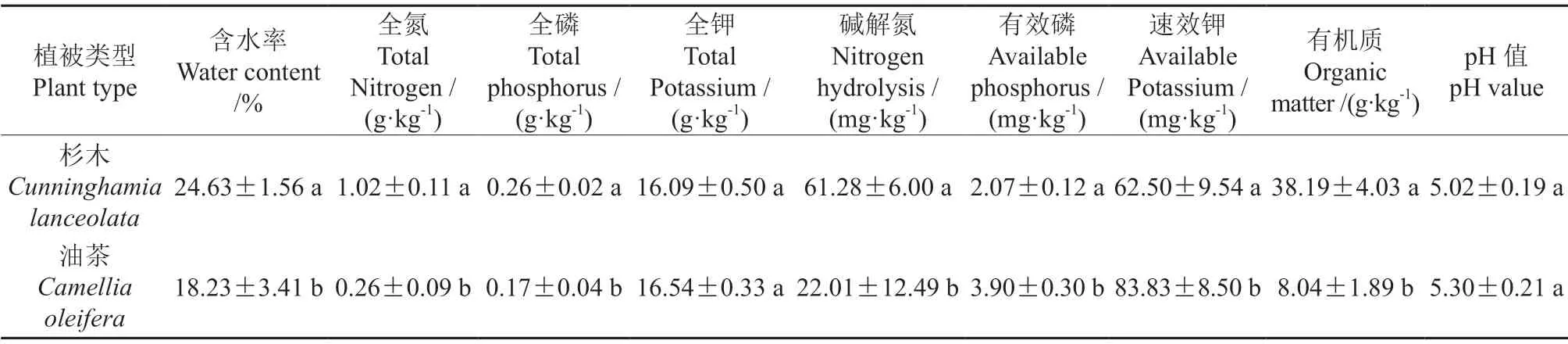
表1 2 种植被类型土壤理化性质†Table 1 Soil physicochemical properties of the 2 plantations
3.2 土壤真菌α 群落多样性
6 个土样中有效系列为309 733 条,有效碱基数为118 113 323 bp,序列平均长度为381 bp,2种林地的Coverage 指数均为99.98%,表明测序结果全面地反映真菌群落在所测土壤样本中的存在状况。2 种林地土壤真菌多样性结果见表2。杉木林的Shannon 指数(3.02)、Ace 指数(170.31)均高于油茶林,说明杉木林土壤中真菌多样性和丰度较高,但是均不存在显著差异(P>0.05)。杉木林的Simpson 指数(0.106 6)低于油茶林,差异不显著(P>0.05),杉木林的Chao1 指数(173.12)显著高于油茶林。2 种植被类型土壤真菌群落的Ace 指数和Chao1 指数均较低,说明2种植被类型中真菌数量较低。

表2 2 种植被类型土壤真菌多样性指数Table 2 Diversity indices of soil fungal communities in the 2 plantations
3.3 真菌群落组成分析
2 种植被类型的土样中共检测出真菌门18个,真菌纲46 个,真菌目74 个,真菌科92 个,真菌属98 个,真菌种116 个,OTU252 个。杉木林和油茶林土壤中相对丰度在1%以上的优势菌门有3 个(图1a),分别为子囊菌门(Ascomycota)、担子菌门(Basidiomycota)和毛霉菌门(Mucoromycota)。杉木林土壤中子囊菌门相对丰度(67.19%)低于油茶林(76.53%),杉木林担子菌门(24.38%)高于油茶林(15.45%),杉木林和油茶林毛霉菌门的相对丰度分别为4.38%和4.94%,3 个优势菌门在2 种植被类型间均不存在显著差异(P>0.05)。
如图1b 所示,2 种植被类型的土壤真菌群落在真菌纲水平上的组成不同。杉木林土壤中相对丰度在1%以上的且鉴定出的优势菌纲有8 个,古根菌纲(Archaeorhizomycetes)的相对丰度最大(28.53%),其次为伞菌纲(Agaricomycetes)(15.13%)、散囊菌纲(Eurotiomycetes)(14.79%)、粪壳菌纲(Sordariomycetes)(9.61%)、银耳纲(Tremellomycetes)(6.75%)、盘菌纲(Pezizomycetes)(4.5%)、锤舌菌纲(Leotiomycetes)(3.72%)、座囊菌纲(Glomeromycetes)(1.85%)。油茶林土壤中鉴定出的相对丰度在1%以上的优势菌纲有7个,相对丰度最大的是散囊菌纲(23.41%),其次为粪壳菌纲(22.52%),古根菌纲(20.4%)、伞菌纲(13.96%)、锤舌菌纲(4.21%)、球囊菌纲(2.91%)、银耳纲(1.35%)。油茶林中散囊菌纲显著高于杉木林(P<0.05),其余优势菌纲在2 种植被类型间差异不显著(P>0.05)。
图1c 所示杉木林和油茶林土壤真菌在属水平上识别出且相对丰度1%以上的属共有9 个,其中7 个属于子囊菌门,分别为Archaeorhizomyces(28.54%、20.4%)、篮状菌属(Talaromces)(11.76%、21.36%)、木霉菌属(Trichoderma)(4.21%、8.11%)、毛壳菌属(Chaetomium)(1.6%、6.18%)、Meliniomyces(2.6%、1.98%)、稻瘟病菌(Magnaporthe)(0.29%,3.08%)和瓶霉属(Phialophora)(0.45%,1.17%)。而粗糙孔菌属(Trechispora)(0.34%,1.96%)和红菇属(Russula)(1.48%,0.04%)则属于担子菌门。油茶林土壤中毛壳菌属和稻瘟病菌显著高于杉木林(P<0.05),Archaeorhizomyces、篮状菌属、木霉菌属、Meliniomyces粗糙孔菌属、瓶霉属和红菇属在2 种植被类型土壤中差异不显著(P>0.05)。

图1 真菌门、纲和属水平上的相对丰度Fig.1 The relative abundances of dominant fungi at the phylum,class and genus levels
3.4 环境因子对土壤真菌群落结构的影响
土壤真菌与土壤理化性质的冗余分析如图2所示。由图2可知,第一排序轴和第二排序轴的RDA 相关分析可知,第一排序轴的解释率为82.89%,第二排序轴的解释率为14.02%,,两个排序轴的解释率为96.91%,说明土壤理化性质对土壤真菌群落结构有较大影响。全磷和速效钾对第一排序轴贡献较大,速效钾对第二排序轴的贡献最大,其次为全磷,综合来看,全磷对真菌群落结构的影响最大,其次为碱解氮。

图2 土壤真菌与土壤理化性质的冗余分析Fig.2 Redundancy analysis(RDA)of the relationships between soil physicochemical properties and fungal communities of A B treatments
优势菌门和优势菌纲对土壤理化性质的响应表3。担子菌门与全磷显著正相关(P<0.05)。粪壳菌纲与碱解氮极显著负相关(P<0.01),与有效磷显著正相关(P<0.05)。银耳纲与全磷和碱解氮极显著正相关(P<0.01),与全氮显著正相关(P<0.05)。锤舌菌纲与全钾显著负相关(P<0.05),unclassified_p_Ascomycota 与全钾极显著负相关(P<0.01)。座囊菌纲与碱解氮和有机质显著负相关(P<0.05)。

表3 土壤真菌优势门(门、纲水平)相对丰度和土壤理化性质相关关系†Table 3 Correlation between soil physicochemical properties and dominant fungal phyla and classes
4 讨 论
4.1 2 种植被类型对真菌优势门、纲和属类群的影响
研究结果表明,杉木林和油茶林的土壤中优势菌门、优势菌纲和优势菌属的组成存在一定的相似,但优势菌门、优势菌纲和优势菌属在2 种植被类型土壤中的相对丰度又有一些差异,这是因为不同植物根系的分泌物和枯落物对菌群多样性和结构有影响[24-26]。杉木林和油茶林土壤中主要优势菌门为子囊菌门和担子菌门,两个菌门在土壤中的丰度之和为91.57%~91.97%。结果与陈秀波[27]、曹红雨[28]、杨立宾[29]等研究结果相似。子囊菌门和担子菌门属于高等陆生真菌,在透气性好的酸性土壤中更容易生长。子囊菌门以腐生营养型菌居多,可以将林分中较难降解的角质素等进行分解,对森林生态系统养分循环具有重要作用[30-31]。油茶林土壤中的子囊菌门高于杉木林,有利于油茶林枯落物的分解,担子菌门对木纤维素的降解、植物菌根的形成等具有促进作用[32],还可以降低病原菌对植物的危害[33-34]。油茶林土壤中担子菌门的相对丰度比杉木林显著下降,说明油茶林发生真菌病害的概率有所增加。在真菌纲水平上,2 种植被类型的群落组成不同,杉木林土壤已鉴定出的优势菌纲的排序为古根菌纲>伞菌纲>散囊菌纲>粪壳菌纲>银耳纲>盘菌纲>锤舌菌纲>座囊菌纲。油茶林的排序为散囊菌纲>粪壳菌纲>古根菌纲>伞菌纲>锤舌菌纲>球囊菌纲>银耳纲。2 种植被类型土壤中识别的优势菌属为Archaeorhizomyces、篮状菌属、木霉菌属、毛壳菌属、Meliniomyces、稻瘟病菌、粗糙孔菌属、瓶霉属和红菇属。与前人研究中青霉属为主要优势菌属略有不同[35],本研究的主要优势菌属为Archaeorhizomyces和篮状菌属,相对丰度之和为40.3%~41.76%,这可能由于林地类型,树种[36-37]、土壤理化性质[38]等不同引起土壤群落结构的变化[39]。
4.2 土壤理化性质对土壤真菌群落及多样性的影响
通过对2 种植被类型土壤真菌的a 多样性分析,杉木林的真菌多样性和丰度均高于油茶林。在纲水平上,真菌群落组成不同,说明杉木林转为油茶林改变了土壤真菌群落结构。土壤的含水率、pH 等因素可单独或相互作用于土壤真菌群落结构[40]。由冗余分析可知,全磷和碱解氮是影响土壤真菌群落结构的主要因素。Lauber 等[41]研究表明子囊菌门与土壤中的氮含量呈显著正相关,本研究中子囊菌门与全氮呈负相关,但不显著。全磷对担子菌门呈显著正相关。碱解氮与3 个菌纲成显著相关(粪壳菌纲、银耳纲、座囊菌纲)。有效磷与2个菌纲呈显著相关(粪壳菌纲、银耳纲)。银耳纲与全氮和全磷呈显著正相关,锤舌菌纲与全钾呈显著负相关。结果说明不同的理化因子对特定的菌纲有显著作用,而土壤中环境因子的改变对真菌门、纲等相对丰度影响较大,进而改变土壤真菌的群落结构组成。
5 结 论
1)杉木林的真菌多样性高于油茶林,但差异不显著。杉木林的真菌丰度显著高于油茶林。全磷和碱解氮是影响真菌群落的主要因子。
2)2 种植被类型土壤中共检测出真菌门18个,真菌纲46 个,真菌目74 个,真菌科92 个,真菌属98 个,真菌种116 个,OTU252 个。杉木林转为油茶林后,子囊菌门和担子菌门是占绝对优势的2 个菌门,担子菌门相对丰度杉木林显著高于油茶。杉木林的优势菌纲古根菌纲、伞菌纲和散囊菌纲。油茶林的优势菌纲为散囊菌纲、粪壳菌纲、古根菌纲。2 种植被土壤中的优势菌属为Archaeorhizomyces和篮状菌属。本研究杉木林转为油茶林后真菌群落结构组成相似,但只研究了杉木转为油茶林后真菌群落结构的变化,随着油茶林种植年限的增加,接下来的研究中还需进一步研究成熟油茶林真菌群落结构的变化,以期为油茶林的精细化管理提供更多可以参考的依据。
