DNA双链断裂修复相关基因XRCC1、XRCC3、TP53、Rad51多态性与结直肠癌易感性关联的Meta分析
2022-02-14苟怡凡周思佳张永吉宁宁康思涵逯晓波
苟怡凡,周思佳,张永吉,宁宁,康思涵,逯晓波
(中国医科大学公共卫生学院卫生毒理学教研室,辽宁 沈阳110122)
结直肠癌(colorectal cancer,CRC)作为常见的恶性肿瘤,近年来在我国的发病率及死亡率均呈上升趋势,2015年我国CRC发病率居发病瘤谱第4位,病死率居死亡瘤谱第5位[1]。研究表明,90%CRC的发生是环境因素与遗传因素交互作用的结果[2]。DNA损伤是常见的遗传因素,DNA损伤修复可以逆转、修复DNA损伤,使其恢复结构,维持遗传物质的稳定性。大量证据提示DNA损伤修复能力的高低与CRC发生风险之间存在关联[3],DNA损伤修复系统缺陷或效率降低可以增加CRC发病风险。常见的DNA修复系统有很多种,错配修复与遗传性CRC发病风险的关联最为密切,但是DNA双链断裂是一种最为严重的DNA损伤,其相应修复系统与CRC发生风险之间的关联目前尚无准确定论。本研究将对DNA双链断裂修复系统基因XRCC1、XRCC3、TP53、Rad51与CRC发病风险的关联进行Meta分析。
XRCC1位于人19号染色体19q13.2[4],是一种参与单链断裂修复(SSBRs)和碱基切除修复(BER)的DNA修复基因[5]。XRCC1蛋白可以修复辐射诱导所引起的损伤[6]。目前发现XRCC1的第194、280、399号有3个多态性位点,其中尤以399位密码子值得引起重视,它位于多聚ADP核糖聚合酶PARP的结合域BRCT-1上,而许多有BRCT-1域的蛋白质参与细胞周期和DNA损伤修复[6]。因此我们选取XRCC1 Arg399Gln进行本次研究。
XRCC3位于人染色体14q32.3,该基因编码RecA/Rad51相关蛋白家族成员,参与同源重组以修复DNA损伤[7]。其最常见的多态性位点是第7外显子上C18067T的序列变异,导致第241位密码子编码的氨基酸变化(Thr241Met),可能影响酶的功能以及与其他涉及DNA损伤和修复的蛋白的相互作用[6,8],因此我们选取XRCC3 Thr241Met进行本次研究。
TP53位于人类染色体17q3.1上,是一种抑癌基因,由11个外显子组成,它编码一种含有393个氨基酸的53 KD蛋白,该蛋白在G1阶段可以被DNA损伤和细胞周期停止的信号磷酸化和激活,从而促使DNA修复或细胞凋亡[9]。TP53中的单核苷酸多态性可能会完全破坏蛋白质的功能,使细胞生长速率异常增高,从而导致大量癌症发生[10]。据报道,TP53 Arg72Pro在癌症的易感性和进展过程中起重要作用[11]。因此我们选取TP53 Arg72pro进行本次研究。
Rad51位于人染色体15q15.1。Rad51可以编码对重组修复双链DNA断裂很重要的蛋白质[12],在HR通路中起着至关重要的作用[13]。到目前为止,已经在Rad51中发现了多个单核苷酸多态性,如135G>C和172G>T。它们位于Rad51启动子的调控元件中,可能与mRNA的表达有关[14]。Rad51 135G>C和172G>T多态性被认为是多种癌症的危险因素[14]。其中,Rad51 135G>C更为常见,Rad51 135G>C多态性在5′UTR区域的定位表明,该多态性可能与mRNA的稳定性和翻译有关[15]。因此我们选取Rad51 135G>C进行本次研究。
目前XRCC1 Arg399Gln、XRCC3 Thr241Met、TP53 Arg72Pro、Rad51 135G>C位点单核苷酸多态性与CRC易感性关系的结论具有不一致性,因此我们进行Meta分析,来综合评价这几种DNA双链断裂损伤修复基因多态性与CRC遗传易感性的关系。
1 资料与方法
1.1 文献检索 在PubMed、中国知网、万方及维普等数据库中搜索,检索跨度时期为建库至2020年9月15日,以“XRCC1(XRCC3,TP53,Rad51)”、“polymorphism”或“多态性”或“SNP”、“colorectal cancer”或“结直肠癌”为检索词。
1.2 纳入标准与排除标准
l.2.l 纳入标准 (1)XRCC1的Arg399Gln(XRCC3的Thr241Met、TP53的Arg72Pro、Rad51的135G>C)位点基因多态性与CRC易感性相关关系的研究;(2)以论著形式发表的病例-对照研究;(3)样本量明确,数据完整,可以获得等位基因频率和(或)基因型频率的数据。
l.2.2 排除标准(1)重复的文献;(2)综述、报道、会议摘要等;(3)无对照组研究的文献;(4)信息不完整的文献。
1.3 资料提取与质量评价 根据Newcastle Ottawa scale(NOS)标准[16],对所有文献进行评价和筛选。由2位参与者独立进行,提取国家、种族、基因检测方法、肿瘤组和对照组的样本总数和基因型分布信息,若有不同意见则请第三人做出评判。文献评分最高分9分,最低分0分;只有5分或以上的高质量文献才会被纳入此次研究。
1.4 统计学方法(1)异质性检验和Meta分析:采用Review Manager 5.3及Stata 16.0进行数据分析,同时对纳入的研究进行异质性检验。根据I2的大小评估异质性强弱,若I2≥50%,说明异质性较强,采用随机效应模型;否则采用固定效应模型。计算OR值及95%可信区间(95%CI)。(2)敏感性分析:逐篇剔除文献并分析。(3)发表偏倚分析:采用Stata 16.0进行Egger′s检验,评估发表偏倚。
2 结果
2.1 文献基本信息 通过文献检索,XRCC1、XRCC3、TP53、Rad51分 别 查 找 到134、42、135、19篇相关文献,通过阅读摘要和全文对文献进行筛选,最终分别纳入11(5篇中文,6篇英文)、15(15篇英文)、8(8篇英文)、6(1篇中文和5篇英文)篇文献。XRCC1累计病例2 448例,对照3 681例;XRCC3累计病例6 150例,对照7 773例;TP53累计病例3 970例,对照4 024例;Rad51累计病例853例,对照800例。XRCC1、XRCC3、TP53、Rad51纳入文献的基本信息及方法学质量评价见表1~4。

表1 纳入XRCC1相关研究文献的基本信息及其质量评价
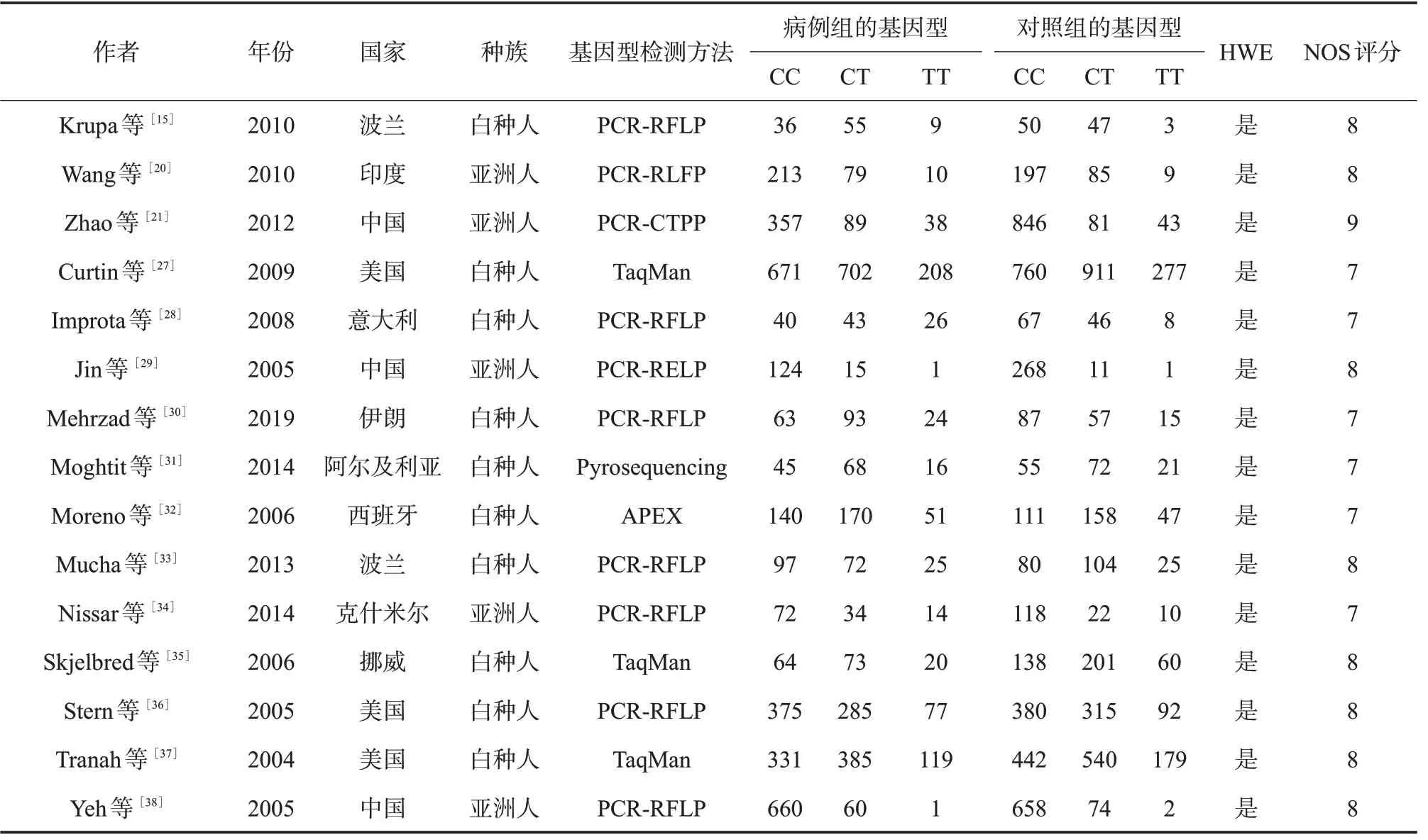
表2 纳入XRCC3相关研究文献的基本信息及其质量评价
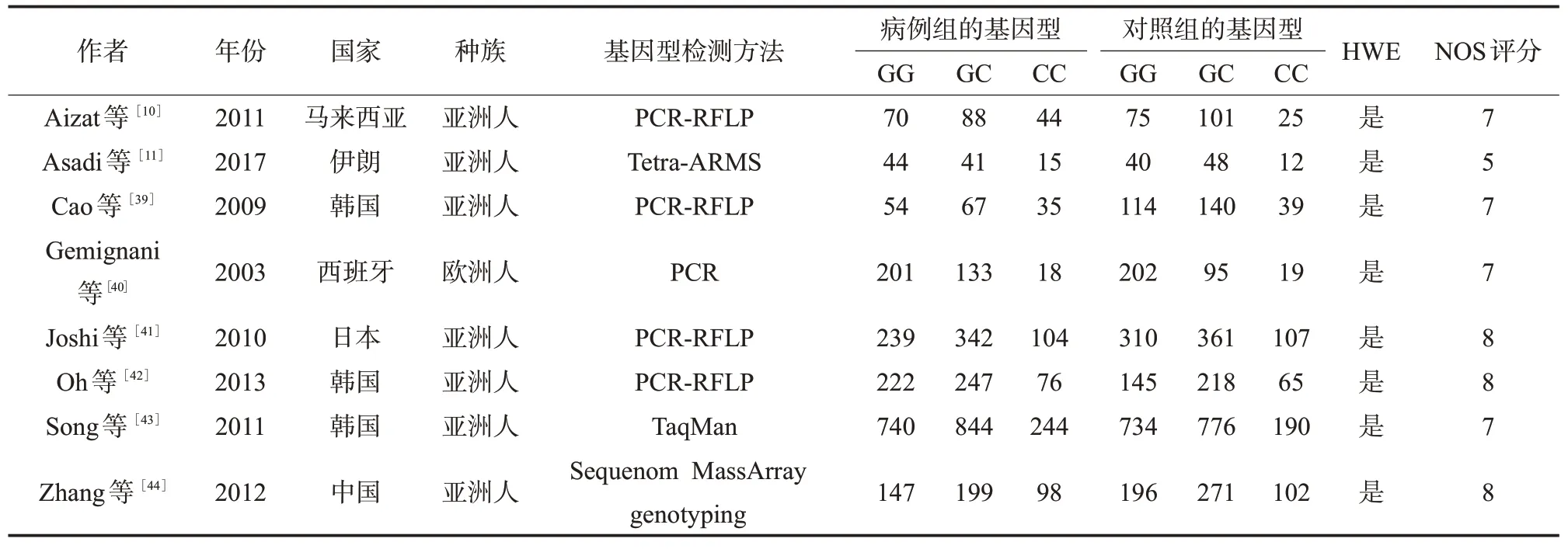
表3 纳入TP53相关研究文献的基本信息及其质量评价
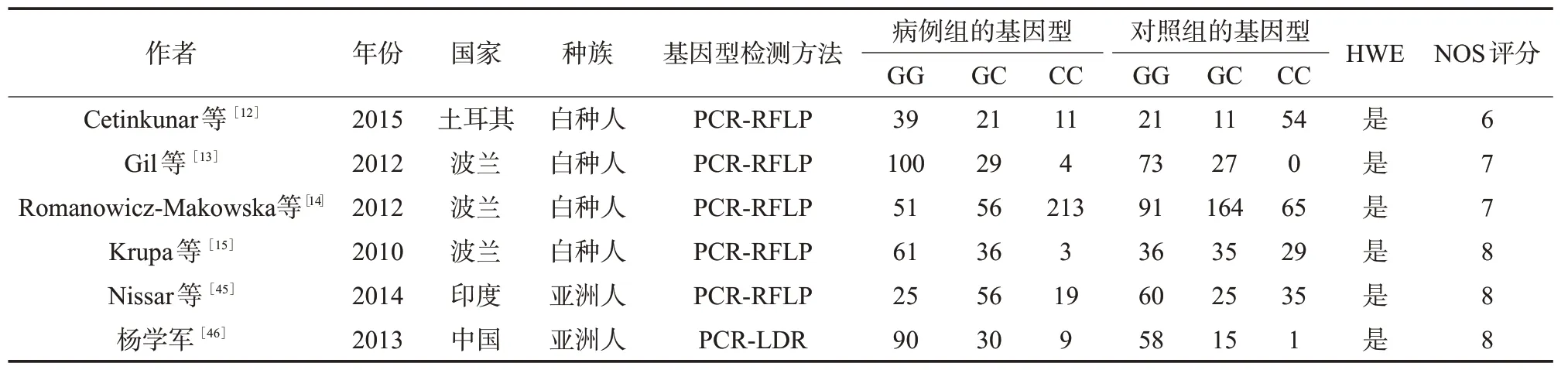
表4 纳入Rad51相关研究文献的基本信息及其质量评价
2.2 异质性检验与Meta分析XRCC1、XRCC3、TP53、Rad51异质性检验采用I2检验。(1)XRCC1(AG+AA/GG):I2=87%,P<0.01。提示各合并的研究数据有显著异质性,采用随机效应模型。Meta分析结果表明,XRCC1 Arg339Gln基因多态性与CRC发病风险无显著相关性(AG+AA/GG)OR=0.98,95%CI:0.72~1.34,P=0.90,见图1。(2)XRCC3(CT+TT/CC):I2=86%,P<0.01。提示各合并的研究数据有显著异质性,采用随机效应模型。Meta分析结果表明XRCC3 Thr241Met基因多态性与CRC发病风险无显著相关性(CT+TT/CC)OR=1.20,95%CI:0.97~1.49,P=0.10,见图2。(3)TP53(CC+GC/GG):I2=39%,P=0.12。提示各合并的研究数据无显著异质性,采用固定效应模型。Meta分析结果表明TP53 Arg72Pro基因多态性与CRC发病风险无显著相关性(CC+GC/GG)OR=1.08,95%CI:0.99~1.18,P=0.10,见 图3。(4)Rad51(CC+GC/GG):I2=91%,P<0.01。提示各合并的研究数据有显著异质性,采用随机效应模型。Meta分析结果表明Rad51 135G>C基因多态性与CRC发病风险无显著相关性(CC+GC/GG)OR=0.98,95%CI:0.46~2.11,P=0.97,见图4。
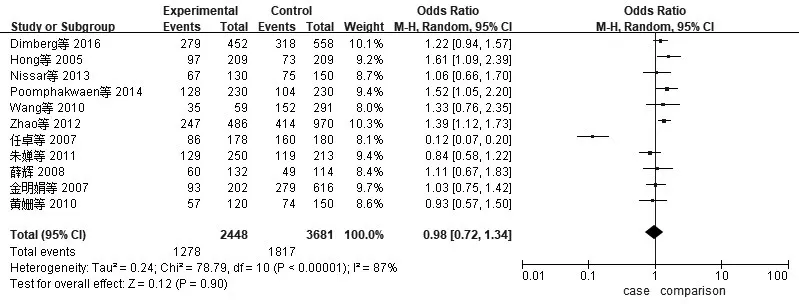
图1 纳入XRCC1相关研究CRC与显性模型AG+AA/GG的森林图
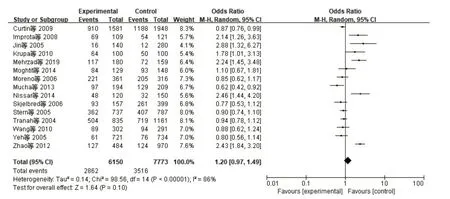
图2 纳入XRCC3相关研究CRC与显性模型CT+TT/CC的森林图
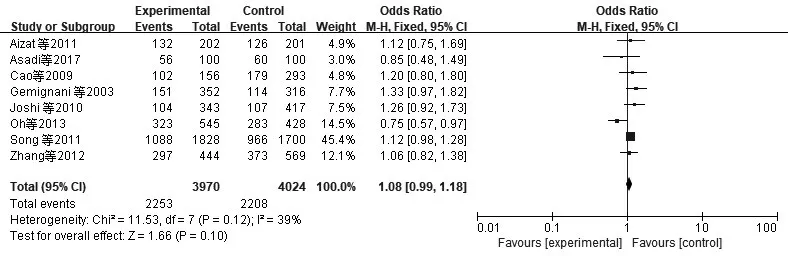
图3 纳入TP53相关研究CRC与显性模型CC+GC/GG的森林图
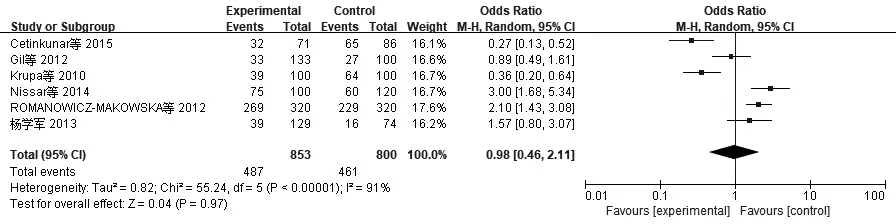
图4 纳入Rad51相关研究CRC与显性模型CC+GC/GG的森林图
2.3亚组分析结果(1)XRCC3:由于种族及环境因素可能会对基因多态性产生影响,因此按本文种族将文献划分并进行亚组分析,其结果表明,XRCC3 Thr241Met基因多态性在亚洲人和白种人中均无意义,见图5。(2)Rad51:按本文种族将文献划分并进行亚组分析,由于亚洲人相关研究数据太少,所以仅对白种人相关研究进行亚组分析,其结果表明,Rad51 135G>C基因多态性在白种人中无意义,见图6。
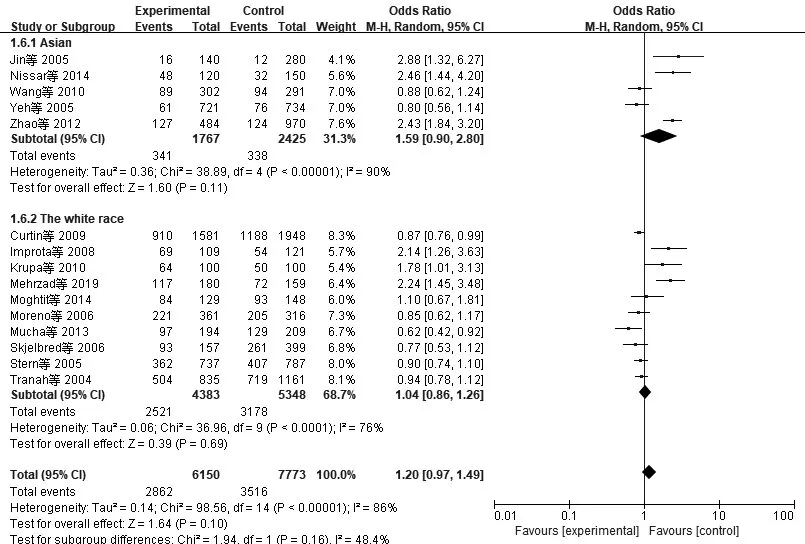
图5 纳入XRCC3相关研究CRC与显性模型CT+TT/CC亚组分析的森林图
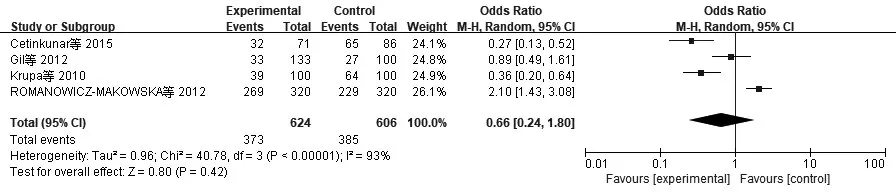
图6 纳入Rad51相关研究CRC与显性模型CC+GC/GG亚组分析的森林图
2.4 敏感性分析 (1)XRCC1:逐一排除分析后,发现剔除任卓等[22]的研究后对OR值产生显著影响(固定效应模型下OR=1.22,95%CI:1.09~1.36,P=0.004),由原来的无关因素变为了危害因素。因此,推测该研究可能是产生异质性的原因之一。(2)XRCC3:逐一排除分析后,发现剔除Yeh等[38]的研究后对亚洲人亚组的OR值产生显著影响,剔除Mehrzad等[30]的研究后对白种人亚组的OR值产生显著影响。推测这两项研究可能是产生异质性的原因之一。(3)TP53:总体而言,逐一排除分析后,各合并效应量无明显改变。但剔除Oh等[42]的研究后对OR值产生显著影响(固定效应模型OR=1.14,95%CI:1.03~1.25),由无关因素变为危害因素。推测该研究可能是产生异质性的原因之一。(4)Rad51:总体而言,逐一排除分析后,各合并效应量无明显改变。发现剔除Cetinkunar等[12]的研究后对OR值产生显著影响,推测该研究可能是异质性产生的原因之一。
2.5 发表偏倚分析 显性模型下4种基因多态性与CRC易感性相关Meta分析的发表偏倚结果如下:(1)XRCC1:Egger′s线性回归结果为t=-0.80,P=0.4422>0.05,不存在发表偏倚。(2)XRCC3:Egger′s线性回归结果为t=2.45,P=0.0294<0.05,存在发表偏倚。(3)TP53:Egger′s线性回归结果为t=-0.25,P=0.8083>0.05,不存在发表偏倚。(4)Rad51:Egger′s线性回归结果为t=-1.02,P=0.3633>0.05,不存在发表偏倚。
3 讨论
CRC发生的常见外因有物理、化学和生物因素等,常见内因有遗传易感性等[17]。DNA损伤修复可逆转机体内、外环境因素导致的DNA损伤,以维持遗传物质的稳定性,是保持细胞基因组完整性的重要机制。在众多类型的DNA损伤中,由电离辐射引起的DNA双链断裂是最严重的致死性DNA损伤,DNA双链断裂修复的常见机制有同源重组(HR)和非同源末端连接(NHEJ)两种通路。在真核细胞中根据断裂的情况,HR和NHEJ可以通过竞争或合作完成修复[4]。经查阅文献发现XRCC1、XRCC3、TP53、Rad51等基因与DNA双链断裂修复以及碱基切除修复等DNA损伤修复过程密切相关。
XRCC1基因编码蛋白可修复DNA损伤包括氧化损伤、烷基化和甲基化损伤等[21]。XRCC1编码的蛋白质通过与多聚ADP核糖聚合酶(PARP)、APE1、DNA连接酶Ⅲ及DNA多聚酶β等相互作用来参与DNA损伤修复[4]。XRCC1 Arg399Gln多态性位点,位于BRCT-1区域,该域保证细胞在G1和S/G2上发生的单链核苷酸缺损都能够得到修复[18]。因此Arg399Gln多态性表型有可能改变XRCC1蛋白的结构与功能,损害XRCC1蛋白与其他酶蛋白的相互作用改变DNA修复活性,从而增加癌症风险[19]。XRCC3在DNA双链断裂的同源修复途径中十分重要。XRCC3是Rad51家族成员之一,现今已发现多种人类XRCC3与Rad51和/或其他Rad51同系物之间的蛋白质相互作用形式或蛋白质复合物,其中Rad51C/XRCC3复合物对于DNA双链断裂所暴露的单链DNA具有很强的结合能力,对于启动重组修复十分重要。另外该复合物能够促进DNA聚集以利于姐妹染色单体之间或同源染色体之间的同源配对[20]。在DNA修复过程中,XRCC3还可组装或稳定Rad51多聚体,以准备重组修复的必要条件[7]。Tp53基因以多条信号通路,多种调控方式参与DNA修复。它可以通过其下游一系列靶基因p21、gadd45等调控细胞周期,使细胞停滞于G1期、G2期等检测点,从而使受损DNA有足够的时间进行多因子参与的修复过程;也可以与DNA修复因子RPA、PCNA、XPp48基因等相互作用,直接参与DNA修复;还可以通过蛋白-蛋白相互作用参与DNA修复[21]。
同源重组HR作为哺乳动物细胞DNA双链断裂修复的主要途径,依靠完整的同源双链来修复断裂的链,其关键步骤是Rad51蛋白诱导的同源链交换。HR修复双链断裂的过程必须首先进行5′-3′末端切除,即降解DNA末端以产生长的3′-ssDNA尾。然后,RecA/Rad51蛋白家族的一个成员与3′-ssDNA尾结合,启动同源配对,并作为链侵入之后DNA合成的引物[22]。
本文通过Meta分析的方法初步探索该4种基因位点多态性在CRC发生、发展中的作用,为CRC的防治提供基础依据。研究结果表明,在显性遗传模型中,4种基因多态性与CRC易感性之间无明显相关性。亚组分析显示,XRCC3 Thr241Met基因多态性在亚种人和白种人中均无意义,Rad51 135G>C基因多态性在白种人中无意义。敏感性分析显示XRCC1、TP53剔除1篇文献后,基因多态性由无关因素变为有害因素,故其与CRC关系仍需进一步探讨与研究。Egger′s检验表明除XRCC3外其他基因遗传模型中均无发表偏倚,因此本研究结果较为可靠。
本研究仍存在不足:(1)部分研究数据由于无法获得文献全文未能被纳入此次Meta分析;(2)XRCC3的研究存在发表偏倚;(3)对于XRCC1、XRCC3、Rad51,由于所纳入研究的条件限制,无法进行进一步的亚组分析来解决异质性较大的问题。
