不同饲养方式对玫瑰冠鸡快慢肌发育及肉质性状的影响
2021-10-21陈若楠杨朝永
胡 波,陈若楠,杨朝永,孙 杰
(石河子大学动物科技学院,新疆石河子 832000)
目前,我国家禽的养殖方式多种多样,饲养方式对家禽肌纤维发育有着重要影响。玫瑰冠鸡是新疆地方特有鸡种,成年鸡鸡冠、肉髯、耳叶和脸均呈鲜红色,胸肌和腿肌健硕,其肉质紧实鲜美、柔嫩多汁、适应能力和抗逆性强[1]。肌肉由肌纤维组成,肌纤维分为快肌纤维和慢肌纤维,比目鱼肌主要以慢肌纤维为主,趾长伸肌以快肌纤维为主[2]。若慢肌肌纤维比例高,则肉色红润,鲜嫩多汁,风味良好;快肌纤维比例高则肉色发白,肉品质下降[3]。
快肌肌钙蛋白(TNNI2)基因与骨骼肌快肌纤维的收缩、肌纤维特性有关[4];肌球蛋白重链1 中肌球蛋白重链1E(MYH1E)和肌球蛋白重链1B(MYH1B)基因与快肌纤维的肌球蛋白生物合成有关,参与肌肉的收缩和代谢[5-6];快白肌肌球蛋白重链(FWM)在家禽快肌纤维的早期发育起关键作用,慢肌肌球蛋白重链(SM)在家禽不同时期肌肉慢肌纤维中表达,运用肌球蛋白重链分子分型的方法准确可靠,能够反映不同类型肌纤维的状况[7];慢速氧化型肌纤维(MYH7)与肌球蛋白生物合成有关;肌钙蛋白T1(TNNT1)又称慢肌肌钙蛋白T,在慢肌纤维中表达[8],主要功能是使复合物与肌球蛋白结合;过氧化物酶增殖物激活受体(PPARD)基因可以加快组织新陈代谢,对于增加慢肌纤维具有重要作用[9]。本研究通过测定不同饲养方式下玫瑰冠鸡快、慢肌纤维组织学特性和肉品质性状,并运用qRT-PCR方法检测快肌MYH1E、MYH1B、FWM和TNNI2基因及慢肌MYH7、TNNT1、SM和PPARD基因的mRNA表达量,探讨不同饲养方式对玫瑰冠鸡快慢肌生长发育规律及快慢肌相关基因与肌纤维比例、肉品质的相关性,为中国优质肉鸡的培育改良提供参考。
1 材料与方法
1.1 试验动物 本研究选取体重相近的1 日龄玫瑰冠鸡400 只,于石河子大学动物科技学院实验站进行分笼饲养,笼养采用立式笼具,饲养密度为0.06 m3/ 只;散养采用水泥地散养,饲养密度为0.2 m2/只。每天饲喂2 次全价鸡饲料,日粮购自新疆正大饲料有限公司,产品成分分析保证值见表1。鸡群常规免疫,自由采食和饮水。在1、7、21、35、49、63、120 日龄分别采集20 只体重相近玫瑰冠鸡的比目鱼肌和趾长伸肌,公母各10 只。采集的样品一部分放入液氮,另一部分放入10%的甲醛固定液中。

表1 产品成分分析 %
1.2 主要试剂及仪器 TRIZOL®Reagent 试剂盒(Invitrogen公司)、Prime ScriptTMRT reagent Kit with gDNA Eraser(TaKaRa 公 司)、LightCycler®480 SYBR Green I Master(Roche 公司)、单克隆抗骨骼肌球蛋白(FAST/SLOW)(Sigma 公司)、小鼠二步法试剂盒(北京中杉金桥生物技术有限公司)、DAB(北京中杉金桥生物技术有限公司)、Roche96 型荧光定量仪(Roche 公司)、石蜡切片机(LM2235,德国莱卡)、生物显微镜及图像分析仪(OLYMPUS-BX40,日本奥林巴斯)、pH 仪(OPTO-STAR,德国)、肉色仪(OPTO-STAR,德国)、质构仪(TA.XTPlus 物性测试仪,英国)。
1.3 快、慢肌纤维组织学测定 将组织大小修剪为1 cm×1 cm×0.5 cm,依次进行脱水、透明、浸蜡、包埋、切片(垂直肌纤维的延伸方向进行横向切片,厚度为5 µm)、烤片、脱蜡、抗原修复、滴加抑制内源性过氧化物酶、一抗,4℃冰箱孵育过夜、二抗、DAB 显色、复染细胞核、脱色、晾干并封片。
采用OLYMPUS BX40 显微镜拍照,Image Pro Plus 6.0 软件分析。每张切片在10×40 倍的视野下进行观察,随机选取5 个视野,测定其每条肌纤维的直径,并进行人工计数阳性肌纤维个数,最后计算出阳性肌纤维所占的百分比。
1.4 肉品质和肌肉质构特性测定 对120 日龄玫瑰冠鸡进行宰杀,宰杀后用消毒过的手术剪取完整胸肌和腿肌进行肉品质和肌肉质构特性分析。参照胡波等[1]的方法计算肉色、pH 和系水力;水分、蛋白质、肌内脂肪(IMF)参照《食品安全国家标准》(GB/T 5009.3—2016)进行测定。肌肉质构特性测定将样本大小修剪为2 cm×2 cm×1 cm;选用P/75 的圆柱形探头,测前速度为4.00 mm/s,测中速度为2 mm/s,测后速度为2 mm/s,下压比例为75%,探头二次压缩间隔时间为5 s;测指标有硬度、胶黏性、弹性和恢复力。
1.5 总RNA 提取和cDNA 合成 按照TRIZOL®Reagent试剂盒的说明书进行RNA 提取,运用超微分光光度计进行RNA 纯度(OD260/280为1.8~2.0)和浓度检测。采用PrimeScriptTMRT reagent Kit with gDNA Eraser 试剂盒进行cDNA 的合成。
1.6 引物设计与合成 根据GenBank 已经公布鸡MYH1E、FWM、TNNI2、MYH1B、MYH7、SM、TNNT1、PPARD和GAPDH基因的序列,采用Primer 5.0 软件进行在线引物设计,引物序列见表2,由上海生工生物工程有限公司合成。

表2 引物信息序列
1.7 实时荧光定量PCR 实时荧光定量反应总体系为20 μL:2×SYBR Green Master Mix 10μL,Primer Forward(10 μmol/μL)1μL,Primer Reverse(10 μmol/μL)1 μL,cDNA1 μL,ddH2O 7 μL;反应程序:95℃预变性5 min,95℃变性10 s,最适退火温度20 s,72℃延伸10 s,45 个循环;最后95℃5 s,65℃1 min,97℃1 s,4℃保存,用于做熔解曲线分析。
1.8 统计分析 将所有的数据用Excel 软件进行统计,结果用平均数±标准差表示。荧光定量PCR 结果使用2-ΔΔCT的方法进行计算。采用SPSS 22.0 软件进行单因素方差分析或独立样本T 检验,Duancan's 法进行显著性检验,相关系数Pearson 法进行相关性分析。P<0.01表示差异极显著,P<0.05 表示差异显著,P>0.05 表示差异不显著。
2 结果与分析
2.1 玫瑰冠鸡快、慢肌纤维免疫组化染色结果 如图1所示,DAB 显色剂会使相应的抗原呈棕色或棕黄色,苏木素染色使细胞核呈蓝色。骨骼肌免疫组织化学染色阳性切片背景均是无色或浅蓝色。快肌抗体对骨骼肌快肌纤维具有特异性的免疫反应。随着日龄增长,快肌纤维形态逐渐增大,且散养组的快肌纤维形态增大快于笼养组。慢肌抗体对骨骼肌慢肌纤维具有特异性的免疫反应。随着日龄增长,慢肌纤维形态逐渐增大,且散养组的慢肌纤维形态增大快于笼养组。

图1 不同饲养方式下玫瑰冠鸡快、慢肌免疫组化染色结果(x40)
2.2 不同饲养方式下玫瑰冠鸡快、慢肌纤维直径比较由表3 可知,随着日龄增长,玫瑰冠鸡快、慢肌纤维直径呈增加趋势,在1、7、21、35、49、63 和120 日龄间存在极显著差异。散养组在35、49、63 和120 日龄时的快肌纤维直径均极显著大于笼养组。散养组35 日龄的慢肌纤维直径显著大于笼养组,49、63 和120 日龄慢肌纤维直径极显著大于笼养组。

表3 不同饲养方式下玫瑰冠鸡的快、慢肌纤维直径 μm
2.3 不同饲养方式下玫瑰冠鸡快、慢肌纤维比例分析由表4 可知,随着日龄增加,快肌纤维比例逐渐增加,120 日龄与1、7 和21 日龄间存在极显著差异;慢肌纤维比例先下降后上升,散养组120 日龄极显著高于7、21 和35 日龄,笼养组120 日龄极显著高于35 日龄,且显著高于21 日龄。相同日龄,快肌纤维比例在63 和120 日龄散养组极显著低于笼养组;慢肌纤维比例在63和120 日龄散养组显著高于笼养组。

表4 不同饲养方式下玫瑰冠鸡快、慢肌纤维比例分析 %
2.4 不同饲养方式下玫瑰冠鸡的肉品质及肌肉质构特性由表5 可知,散养组胸肌的粗蛋白质、系水力极显著高于笼养组,而粗脂肪极显著低于笼养组,pH 和水分显著低于笼养组。散养组腿肌的粗蛋白质极显著高于笼养组,系水力显著高于笼养组,水分和粗脂肪显著低于笼养组。由表6 可知,散养组胸肌的弹性显著高于笼养组;散养组腿肌的胶黏性极显著高于笼养组,硬度、弹性和恢复力显著高于笼养组。
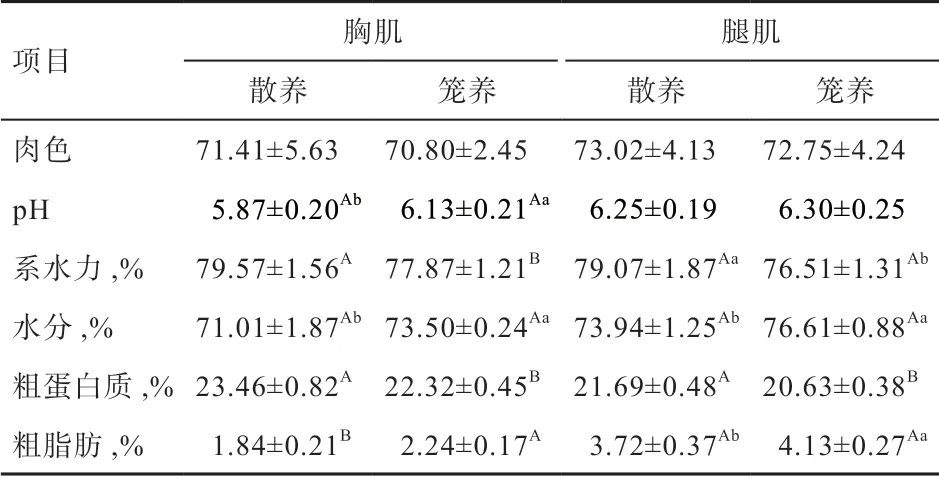
表5 不同饲养方式下玫瑰冠鸡肉品质
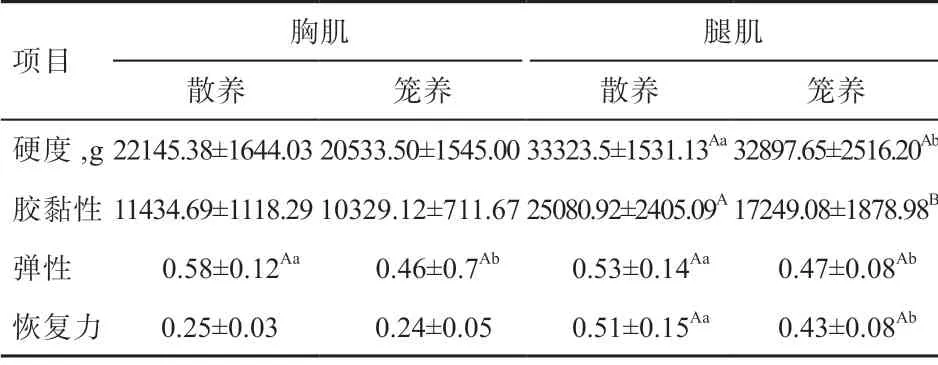
表6 不同饲养方式下玫瑰冠鸡的肌肉质构特性
2.5 快肌MYH1E、MYH1B、FWM和TNNI2基因的mRNA表达量分析 由图2 可知,散养组MYH1E、MYH1B、FWM和TNNI2基因mRNA表达量在1、21、63 和120 日龄均小于笼养组。图2-A 中,散养组MYH1E基因的mRNA 表达量在49 日龄显著低于笼养组;图2-B中,散养组MYH1B基因的mRNA 表达量在21、120日龄极显著低于笼养组;图2-C 中,散养组FWM基因的mRNA 表达量在120 日龄极显著低于笼养组,35、63 日龄显著低于笼养组;图2-D 中,散养组TNNI2基因表达量在63 日龄显著低于笼养组,120 日龄极显著低于笼养组。

图2 不同日龄玫瑰冠鸡趾长伸肌中MYH1E、MYH1B、FWM 和TNNI2 基因的差异表达
2.6 慢肌MYH7、SM、PPARD和TNNT1基因的mRNA表达量分析 由图3 可知,MYH7、SM、PPARD和TNNT1基因mRNA表达量在1、7、21、35、63 和120 日龄散养组均大于笼养组。图3-A 中,散养组MYH7基因的mRNA 表达量在7、35 和63 日龄显著高于笼养组;图3-B 中,散养组SM基因的mRNA 表达量在1 和21 日龄极显著高于笼养组;图3-C 中,散养组PPARD基因的mRNA 表达量在1、7、21 和120 日龄极显著高于笼养组,35 日龄显著高于笼养组;图3-D中,散养组TNNT1基因的mRNA 表达量在21、35 日龄极显著高于笼养组,1 和63 日龄显著高于笼养组。
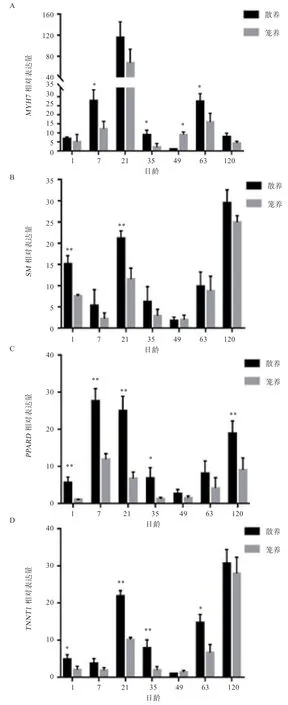
图3 不同日龄玫瑰冠鸡比目鱼肌中MYH7、SM、PPARD 和TNNT1 基因的差异表达
2.7 快、慢肌相关基因表达量与肌纤维比例的相关性 由表7 可见,散养组快肌纤维比例与FWM基因的mRNA 表达量呈正相关(P<0.01),笼养组快肌纤维比例与MYH1E、FWM和TNNI2基因的mRNA 表达量呈正相关(P<0.05)。散养组慢肌纤维比例与MYH7、SM和TNNT1基因的mRNA 表达量呈正相关(P<0.05),笼养组慢肌纤维比例与MYH7、SM基因的mRNA 表达量呈正相关(P<0.05)。
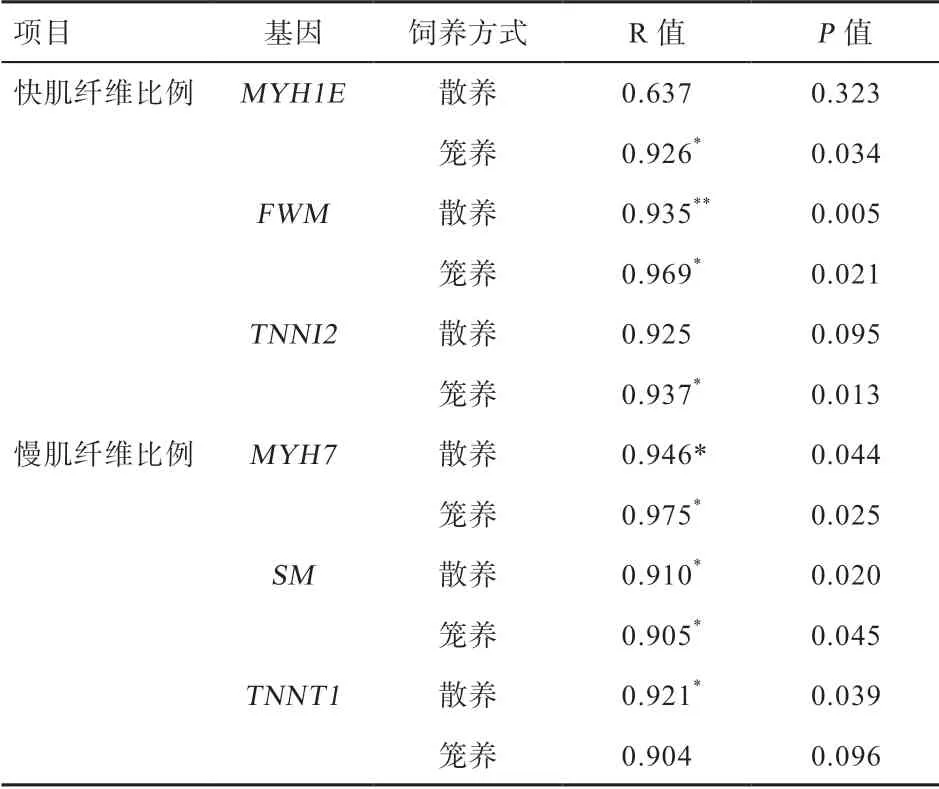
表7 玫瑰冠鸡快、慢肌相关基因表达量与肌纤维比例相关性分析
2.8 快、慢肌相关基因表达量与肉品质性状的相关性由表8 可见,不同饲养方式下,肌肉肉色与MYH7、TNNT1基因的mRNA 表达量均呈正相关(P<0.05),散养组肌肉肉色与SM基因的mRNA 表达量呈正相关(P<0.05),笼养组肌肉肉色与SM基因的mRNA 表达量呈正相关(P<0.01)。笼养组肌肉pH 与MYH1B基因的mRNA 表达量呈正相关(P<0.05),与MYH7(P<0.05)、TNNT1基 因(P<0.01)的mRNA 表 达量呈负相关。散养组肌肉粗蛋白质与TNNT1基因的mRNA 表达量呈正相关(P<0.05),笼养组肌肉粗蛋白质与MYH1E基因的mRNA 表达量呈正相关(P<0.05)。散养组肌肉弹性与MYH1B基因的mRNA 表达量呈负相关(P<0.05),笼养组肌肉弹性与TNNI2基因的mRNA 表达量呈负相关(P<0.05)。

表8 玫瑰冠鸡快慢肌相关基因表达量与部分肉品质相关性分析
3 讨 论
3.1 不同饲养方式对玫瑰冠鸡肌纤维类型发育的影响 畜禽初生时肌肉主要由慢肌纤维组成,出生后随着生长速度加快,肌肉纤维的酵解性增强,慢肌纤维所占比例随年龄的增长而逐渐下降,快肌纤维所占比例逐渐上升[10]。氧化型肌纤维比例会随着周龄增加而降低,酵解型肌纤维比例随着周龄增加而增加[11]。有研究报道,肌纤维直径随着周龄增加而增大,且不同类型的肌纤维直径也有所不同,快肌纤维直径大于慢肌纤维直径[12]。本试验中,玫瑰冠鸡慢肌纤维直径极显著小于快肌纤维直径,且散养组显著大于笼养组,与前人的研究结果一致[12]。
本研究中,随着日龄增长,玫瑰冠鸡趾长伸肌中快肌纤维比例逐渐上升,散养组小于笼养组;比目鱼肌中慢肌纤维比例先下降后上升,散养组大于笼养组,这可能是由于散养组活动量大,经常有跑跳等爆发力动作,导致慢肌纤维比例增加。李江华[13]研究运动对大鼠腓肠肌肌纤维类型百分比构成的影响发现,运动引起II型肌纤维凋亡,促进I 型肌纤维表达,这与本文得出的结果相类似。饲养方式不同会影响肌纤维类型变化,散养可以促进I 型肌纤维的表达[14-15]。快肌纤维含量高的肉质风味一般较差,这是由于其中的不饱和脂肪酸和游离氨基酸等含量较少[16]。肌肉中慢肌纤维所占比例越大,肌肉的感官程度越好,肉的嫩度与色泽也更好[17-18],肌肉品质越好,因此提高慢肌纤维的比例是改善畜禽肉品质的主要途径。研究鸡骨骼肌中快慢肌的调控机制,对于改善鸡肉品质,提升肌肉风味,具有重要意义。
3.2 不同饲养方式对玫瑰冠鸡肉品质性状的影响 肌肉品质主要从肌肉的物理特性、化学成分和质构特性进行评价。肉色、pH 和系水力是评定肌肉品质的物理指标,肉色能反映肌肉品质的新鲜度;pH 与肌糖原的酵解速率有关,是影响肌肉酸味的主要因素[19];系水力代表肌肉的持水能力,影响肌肉的嫩度和多汁性[20]。水分、IMF 和粗蛋白是肌肉中常规化学成分,水分可以直接反映肉的色泽,粗蛋白是决定肉品质的营养成分;IMF 含量越高,肌肉品质越细嫩[21-22]。肌肉硬度体现了肉质的强度、韧性等能力,恢复力、弹性是肌肉压缩过程中回弹的能力[23-24]。
本试验中,散养玫瑰冠鸡有利于提高肉色和系水力、降低pH,肉色提高是由于散养促进肌肉血红蛋白和肌红蛋白含量提高,肌肉中系水力与滴水损失率、蒸煮损失率呈负相关,系水力越高,滴水损失率和蒸煮损失率越低[25-26]。玫瑰冠鸡散养组肌肉水分含量低于笼养组,粗蛋白质含量高于笼养组,表明其营养价值高于笼养组。玫瑰冠鸡散养组腿肌的硬度显著大于笼养组,是由于散养组活动空间大,鸡群之间互相追逐,运动量变大,导致肌肉变得紧实粗壮;其次是肌肉中的慢肌纤维变多,密度增大,导致肌肉硬度变大。刘兴余等[23]通过TPA 法来评定不同品种鸡肌肉品质的差异,得出肌肉的多汁性与质构特性的硬度呈正相关。玫瑰冠鸡肌肉中弹性散养组显著大于笼养组,肌肉的弹性越大,肌肉越有嚼劲、口感越好,这是由于散养提高了慢肌纤维比例和肌纤维密度相对增加所致。
3.3 不同饲养方式对玫瑰冠鸡快慢肌相关基因表达量的影响 趾长伸肌主要是由快肌纤维组成,饲养方式对肌纤维发育有很大影响,散养能够降低快肌纤维的表达[27]。本研究中,散养组玫瑰冠鸡趾长伸肌中MYH1E、MYH1B、FWM和TNNI2基因mRNA 的表达量小于笼养组,表明散养有利于降低趾长伸肌中快肌纤维的表达。120 日龄玫瑰冠鸡MYH1B、FWM基因mRNA 的表达量在趾长伸肌中均较高,推测120 日龄是快肌纤维转化的关键节点,有待进一步研究。
比目鱼肌主要是由慢肌纤维组成,本研究中散养组玫瑰冠鸡比目鱼肌中MYH7、SM、PPARD和TNNT1基因mRNA 的表达量大于笼养组。MYH7、PPARD基因mRNA 表达量随着日龄的增长呈下降趋势,这与张贝贝的研究结果相类似[28]。在家禽生长过程中,由于体重、活动量的需求增加,快肌纤维的需求加大,慢肌需求相对减少,本研究中快肌基因的表达量逐渐上升,慢肌基因的表达量逐渐下降,这与快肌肌纤维比例逐渐上升、慢肌纤维比例逐渐下降相一致。
3.4 快慢肌相关基因表达量与肌纤维比例、肉品质性状的关系 本试验中,MYH7基因mRNA 表达量与慢肌肌纤维比例均呈显著正相关,这与张贝贝[28]试验结果相符。笼养组MYH1E基因mRNA 的表达量与快肌肌纤维比例呈显著正相关,章明等[29]在挖掘清远麻鸡和科宝鸡肌肉差异表达基因中证明了科宝鸡快肌纤维比例高于清远麻鸡,且MYH1E基因在科宝鸡快肌中高表达。
本研究中,MYH7和SM基因mRNA 的表达量与肉色呈显著正相关。MYH7和SM基因作为慢肌肌纤维的标记基因,其与慢肌纤维的比例呈正相关,且慢肌纤维比例与肌肉的色泽、品质密切相关,主要由慢肌中血红蛋白和肌红蛋白含量高所导致[30-31]。笼养组MYH1B基因mRNA 的表达量与pH 呈显著正相关。pH 与肌糖原的酵解速率有关,而MYH1B基因是快肌酵解型的标记基因,是影响肌肉酸味的主要因素,IIb 型肌纤维的数量比例越高,糖酵解能力越强,pH 越低[32]。
散养组TNNT1基因的mRNA 表达量与肌肉硬度达到显著正相关,肌肉的慢肌纤维越多,肌纤维直径越小,肌纤维密度相对就越大,肌肉的硬度也就越大[24]。散养组MYH1B基因的mRNA 表达量与肌肉弹性达到显著负相关,MYH1B基因为快肌纤维的表达基因,与肌肌纤维的构成、收缩有关,而弹性指的是肉质在压缩形变后恢复的程度,弹性又与肌肉的收缩密切相关[33]。
4 结 论
本试验条件下,散养玫瑰冠鸡提高了慢肌纤维比例、系水力、粗蛋白质、弹性和慢肌MYH7、SM、PPARD、TNNT1基因的表达量,降低了快肌纤维比例、水分、粗脂肪和快肌MYH1E、MYH1B、FWM、TNNI2基因的表达量,散养更加有利于提高肌肉品质。
