中部地区家养山羊线粒体遗传多样性与母系起源研究
2021-10-21赵金艳王先宁尹慧茹哈斯通拉嘎王拥庆韩浩园
赵金艳,权 凯,王 俊 ,王先宁,尹慧茹,李 君,哈斯通拉嘎,王拥庆,韩浩园*
(1.河南牧业经济学院动物科技学院,河南郑州 450046;2.沈丘县农牧科技研发中心,河南周口 466300)
山羊属于驯化较早的牲畜之一。山羊群体具有多母系起源,遗传多样性丰富。家养山羊(Capra hircus)由野生山羊进化而来,捻角野山羊(C.aegagrus)和旋角野山羊(C.falconeri)被认为是家养山羊的两大野生祖先。线粒体DNA(Mitochondrial DNA,mtDNA)是动物唯一的核外遗传物质,遵循严格的母系遗传[1]。其中,片段长度为1 220 bp 的D-环控制区(D-loop)受自然选择影响较大,存在巨大变异,所以广泛应用于哺乳动物群体遗传多样性、物种亲缘关系和种群间系统进化等方面的研究[2]。Naderi 等[3]发现7 个高度分化的线粒体单倍型分支(A、B1、B2、C、D、F、G)。
我国拥有丰富的山羊品种资源,长期的自然选择和人工选择可能是导致南北地区家养山羊遗传多样性和种质分化的主要原因。中部地区地方山羊品种丰富,有山西省太行黑山羊、山东省济宁青山羊、河南省伏牛白山羊、槐山羊和尧山白山羊等。其中,太行黑山羊是极其珍贵的食品资源,对饲养环境有较强的适应性,放牧、圈养均能适应,尤其是肉质美味可口,目前由于个体小、生长慢、屠宰率低等原因群体数量急剧减少[4];济宁青山羊是优良的羔皮用山羊品种,具有性成熟早、多胎多羔、常年发情、抗病力强等优良种质特性,近年来,济宁青山羊由于受体型瘦小、生长速度缓慢等缺点的限制,纯种处于濒危状态[5-6];伏牛白山羊具有产肉性能好、屠宰率高、繁殖性能好、耐粗饲、抗病力强、适宜山地放牧等优点;槐山羊具有适应性强、繁殖力强、抗病力强、肉质鲜美、皮张质量好等优点;尧山白山羊是肉皮兼用型山羊品种,具有个体大、抗病力强、四肢粗壮、屠宰率高等特性,尤其是肉质膻味较轻而备受当地消费者青睐,近年来开展肉羊杂交,纯种尧山白山羊分布区域缩小、数量锐减[7]。由于市场竞争、杂化等各种原因,这些品种数量近年来均有下降,为了保护山羊本地品种种质资源的多样性,有必要系统开展中部地区山羊品种群体遗传结构和遗传多态性的研究,为中部地区山羊群体种质的稳定、持续发展奠定理论研究基础。
本研究对中部地区的伏牛白山羊、槐山羊、济宁青山羊、太行黑山羊和尧山白山羊的线粒体D-loop 区遗传多样性、遗传结构及母系起源进行研究,旨在为中国中部地区山羊地方品种资源的评估、保护、利用及山羊品种起源、进化研究等提供科学依据。
1 材料与方法
1.1 样本采集 本研究共采集145 个本地山羊品种外周血液样本,包括20 个伏牛白山羊、74 个槐山羊、28 个济宁青山羊、13 个太行黑山羊、10 个尧山白山羊,采集样本的羊只年龄段为6 月龄至4 岁,随机采集各品种保种场的雌、雄山羊个体,样本采集后置于-20℃保存。
1.2 血液总DNA 提取 采用北京艾德莱生物科技有限公司动物基因组DNA 提取试剂盒提取山羊外周血基因组DNA,利用Thermo Nanodrop 1000 检测OD260/OD280与DNA 浓度,稀释至20 ng/μL,4℃保存备用。
1.3 山羊D-loop 区序列PCR 扩增 山羊线粒体D-loop区上游引物(5'-AACCACTATTAACCACATCTA-3')和下游引物(5'-CACTTACCATGTAAAAGACCC-3')序列由生工生物工程(上海)股份有限公司合成。PCR 扩增体系为25 μL:1 μL DNA 模板、1 μL 上/ 下游引物(10 μmol/L)、9.5 μL ddH2O、12.5 μL TaqPCR Master Mix(北京康为世纪生物科技有限公司)。PCR 扩增程序为:95℃预变性4 min;94℃变性30 s,58 ℃退火30 s,72℃延伸60 s,36 个循环;72℃延伸9 min,4℃保存。利用1%琼脂糖凝胶电泳检测PCR 产物,并由生工生物工程(上海)股份有限公司进行双向测序。
1.4 数据分析 利用Lasergene 软件包SeqMan 软件拼接双向测序的序列,利用MEGA 6.0 软件[8]进行多重比对分析。下载9 条参考序列:旋角野山羊(AB044305)、捻角野山羊(AB004076)、Lineage A(AY155721)、Lineage B(DQ121578)、Lineage C(DQ188892)、Lineage D(AY155952)、Lineage D(DQ188893)、Lineage F(DQ241349)、Lineage G(EF618084),利用MEGA6.0 软件构建NJ 系统发育树,自举检验重复数为1 000,并基于Kimura 2-parameter(K2P)模型[9]计算济宁青山羊、伏牛白山羊、太行黑山羊、槐山羊和尧山白山羊间遗传距离。
利用DnaSP V5 软件进行5 个山羊品种线粒体D-loop 区遗传多态性分析、单倍型分析、品种间遗传分化、基因流分析和中性检验[10],利用NETWORK 10.2.0.0 软件(Fluxus Technology Ltd.,Kiel,Germany)构建济宁青山羊、伏牛白山羊、太行黑山羊、槐山羊和尧山白山羊单倍型网络图。
2 结果与分析
2.1 遗传多态性分析 本研究对槐山羊、太行黑山羊、济宁青山羊、伏牛白山羊和尧山白山羊线粒体D-loop遗传多样性进行分析,总位点数为1 138 bp,缺失位点为156 个,用于计算遗传多样度的位点为982 个(不包括缺失位点),其中,多态位点140 个(排除缺失位点),单态位点842 个。突变位点共定义63 个单倍型,其中,9 个(Hap 3、5、8、13、16、20、26、30、33)为 共享单倍型,品种特异单倍型为54 个。由表1 可知,5 个山羊品种总单倍型多样度(Hd)为0.965±0.004,核苷酸多样度(Pi)为0.013 88±0.000 66,研究发现伏牛白山羊的单倍型多样度最低(0.831),槐山羊的单倍型多样度最高(0.957),太行黑山羊的核苷酸多样度最低(0.011 93),济宁青山羊的核苷酸多样度最高(0.014 34)。
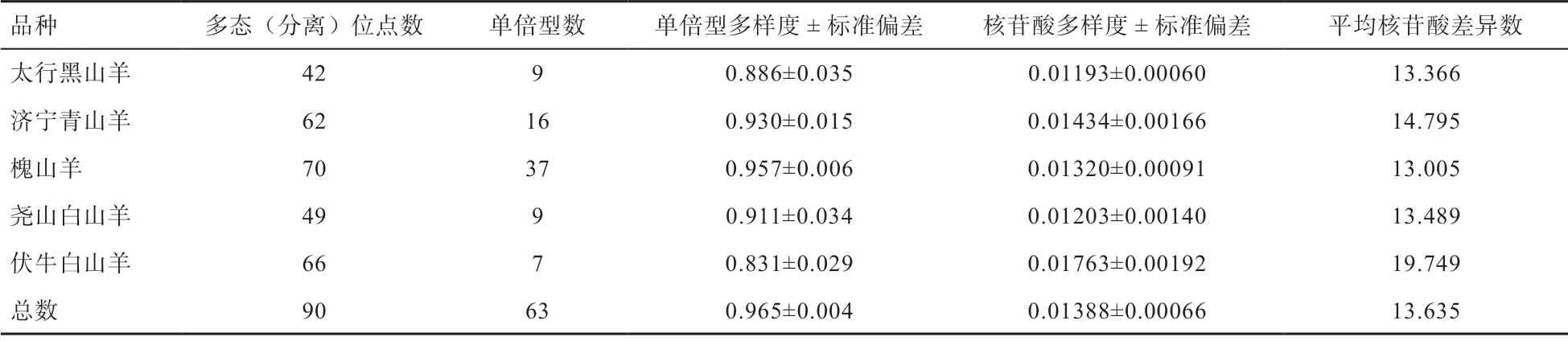
表1 5 个山羊品种线粒体D-loop 遗传多态性分析
2.2 群体结构分析 本研究将捻角野山羊、旋角野山羊和6 个D-loop 分支(Lineage A、B、C、D、F、G)作为参考序列,构建济宁青山羊、太行黑山羊、槐山羊、伏牛白山羊和尧山白山羊系统进化树(图1、2),结果表明5 个山羊品种与捻角野山羊聚类,说明本研究的5 个山羊品种母系起源为捻角野山羊;大部分山羊个体(130 个)属于支系A(Lineage A),极少数个体(15个)属于支系B(Lineage B);支系A 包括济宁青山羊、太行黑山羊、槐山羊、伏牛白山羊和尧山白山羊,支系B 包括济宁青山羊、槐山羊、伏牛白山羊和尧山白山羊。如表2 所示,5 个品种间遗传距离为0.013 6~0.018 8,均未达到亚种间遗传距离。伏牛白山羊与其他4 个品种的亲缘关系相对较远,与济宁青山羊亲缘关系最远,尧山白山羊与其他4 个品种的亲缘关系相对较近。
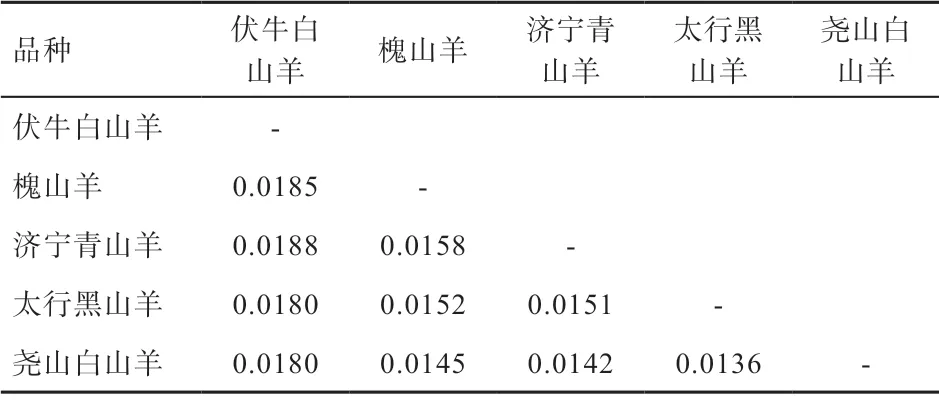
表2 5 个山羊品种间遗传距离
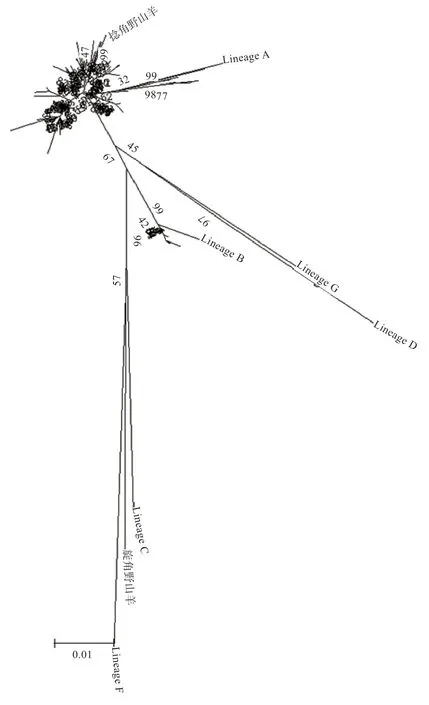
图1 5 个山羊品种系统进化树原始树
2.3 群体遗传分化分析 遗传分化结果如表3 所示,品种间核苷酸平均差异数(Kxy)为11.406 8~16.242 9,遗传分化系数(Fst)为0.016 6~0.169 3,核苷酸歧异度(Dxy)为0.011 6~0.016 5。伏牛白山羊与济宁青山羊间的Kxy和Dxy最大,说明2 个品种间核苷酸差异最大,遗传关系最远,与遗传距离结果一致。伏牛白山羊与尧山白山羊和太行黑山羊间Fst达到明显分化程度,其他品种间均为中度分化或分化程度很弱,槐山羊与济宁青山羊和尧山白山羊、济宁青山羊与尧山白山羊之间均分化程度很弱,说明品种间存在基因交流,与基因流(Nm)为3.92 的结果一致。

表3 品种间遗传分化系数
2.4 群体扩张分析 由表4 可知,中性检验结果表明伏牛白山羊、济宁青山羊和太行黑山羊Fu and Li's F 检验结果为正值,且显著偏离中性,表明伏牛白山羊、济宁青山羊和太行黑山羊近期经历过瓶颈效应和平衡选择。槐山羊和尧山白山羊Fu and Li's F 检验均为P>0.10,不显著,符合动态平衡,未出现群体大规模扩张或瓶颈现象,群体大小保持相对稳定。
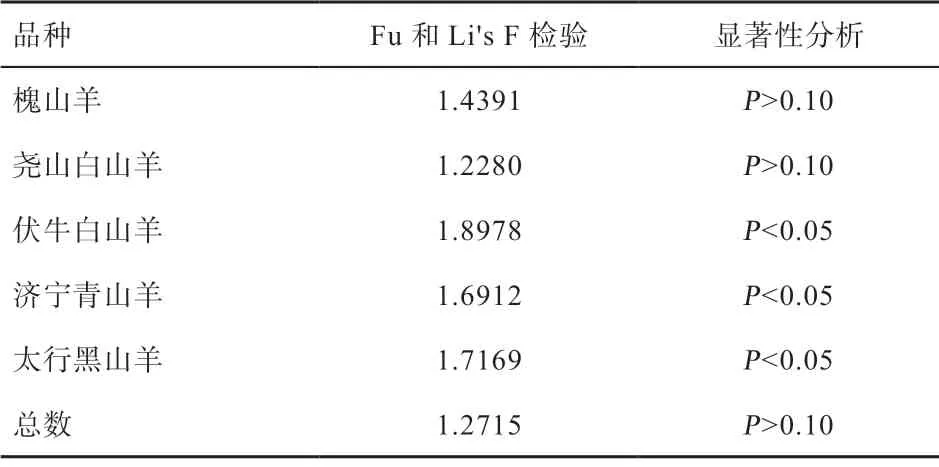
表4 5 个山羊品种群体扩张中性检验结果
2.5 群体单倍型及母系起源分析 如表1 所示,本研究共发现63 个单倍型,共有单倍型为9 个,品种特异单倍型为54 个,槐山羊单倍型最多,为37 个,济宁青山羊定义16 个单倍型,太行黑山羊与尧山白山羊均为9 个单倍型,伏牛白山羊单倍型最少为7 个。由图3 可知,63 个单倍型可分为2 个单倍群(Haplogroup A 和Haplogroup B),分别对应系统发育树中的支系A 和支系B,说明济宁青山羊、槐山羊和伏牛白山羊存在2 个母系起源(支系A 和支系B),尧山白山羊和太行黑山羊为单个母系起源(支系A)。且每个支系呈现“星状”,分布清晰,没有交叉。
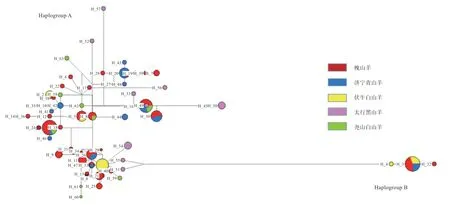
图3 单倍型网络图
3 讨 论
3.1 中部地区山羊遗传多样性Hd和Pi是评价群体遗传多样性和分化的重要指标[11]。本研究发现5 个山羊品 种 总Hd为0.965±0.004,Pi为0.013 88±0.000 66。其中,太行黑山羊Hd为0.886,Pi为0.01193。闫益波等[4]研究发现太行黑山羊Hd值为0.998;武建亮等[12]研究结果为0.800 3。本研究结果位于这2 个数值之间,说明山西太行黑山羊群体多态性较高,近年来群体遗传多样度较为丰富,并维持稳定,具有一定的选择潜力。本研究中伏牛白山羊Hd为0.831,Pi为0.017 63,与李婉涛等[13]人发现的伏牛白山羊群体线粒体D-loop 区存在着丰富的多态性结果一致。济宁青山羊在5 个品种中遗传多样度较高,Hd为0.930,Pi为0.013 20,遗传多样性信息丰富,提供的遗传信息量大,这可能与品种选育过程中育种群体导入了外血、群体选育时间短等因素有关[14]。
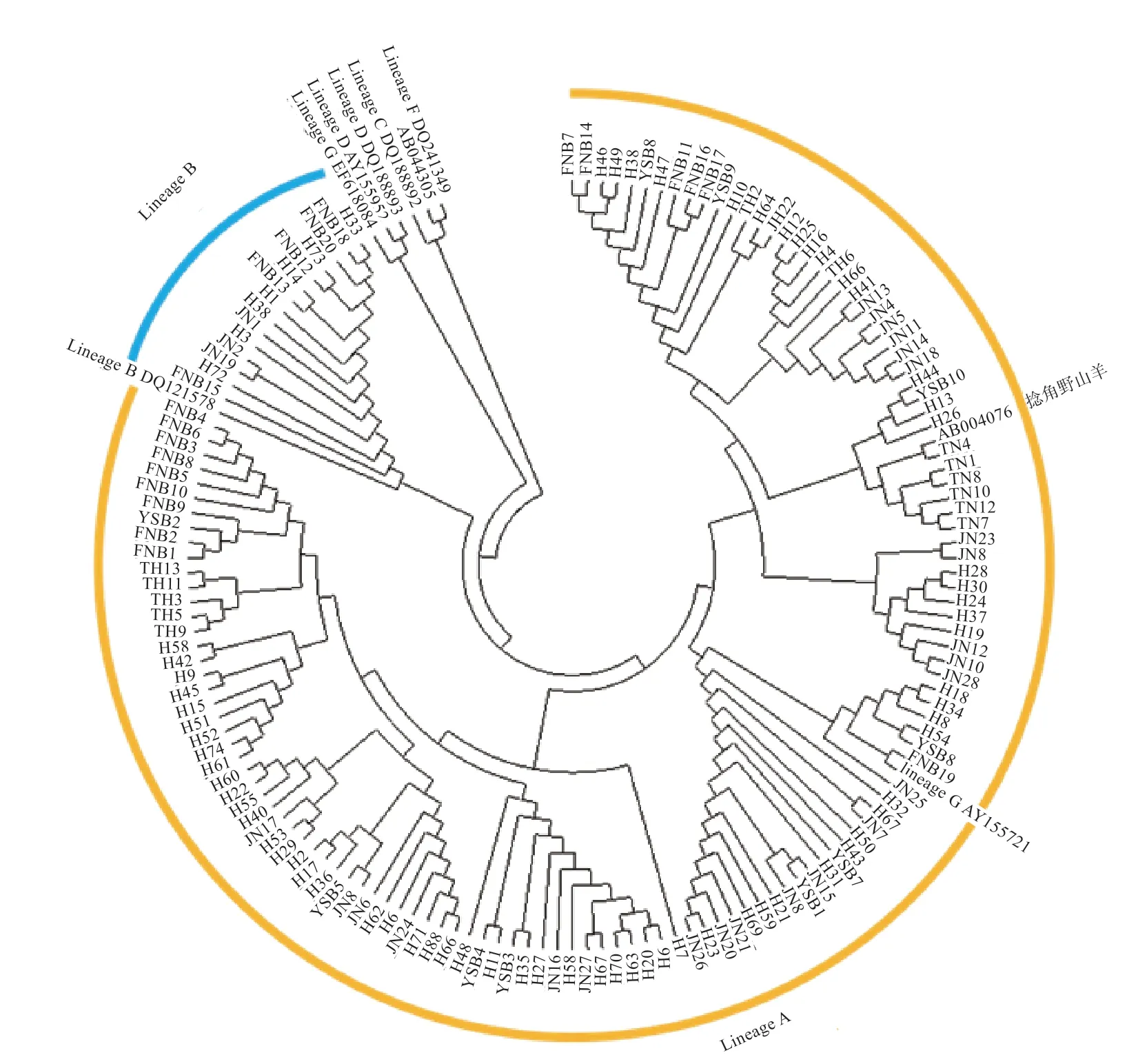
图2 5 个山羊品种系统进化树自展一致树
郝荣超等[15]曾对中国南部地区家养山羊遗传多样性进行研究,发现南部地区家养山羊平均Hd和平均Pi分别为0.954 和0.019 57,西北地区山羊分别为0.919和0.019 08,与本研究中原地区山羊品种遗传多样性相近。有研究针对河南5 个地方山羊品种的遗传多样性进行研究,发现河南省地方山羊的变异主要存在于品种内,品种间的变异相对较小,有足够的遗传变异适应环境的变化和人们的选择需要[16]。
3.2 群体结构分析 本研究遗传距离结果表明5 个品种间遗传距离为0.013 6~0.018 8,郝荣超等[15]发现西北地区山羊品种间遗传距离为0.016~0.019,南部地区品种间遗传距离为0.017~0.025,说明中原地区山羊品种间遗传距离较近,品种间交流较南方和西北地区更频繁。本研究结果显示太行黑山羊与其他4 个品种间的遗传距离范围为0.014 2~0.018 0,均未达到亚种间遗传距离。闫益波等[4]研究表明山西太行黑山羊与槐山羊的遗传距离为0.014,山西太行黑山羊和槐山羊分布的东北和华东地区冬季寒冷,对毛皮性状具有较大的选种要求,这2 个品种都以产绒性状和板皮性状为优,因而群体间起源较近。
Fst可以反映群体近交或群体间遗传分化程度。本研究中除伏牛白山羊与太行黑山羊、尧山白山羊之间达到明显分化,其他品种间Fst为0.016 6~0.122 0,说明群体间的遗传分化水平低,而且基因流大于1,均质化作用强,能够抵制遗传漂变作用,防止群体间遗传分化发生。王可等[6]也发现济宁青山羊的遗传多样性得到了保留,群体间的基因流较大,遗传分化水平较低。闫益波等[4]发现山西太行黑山羊与槐山羊的群体间分化程度较低,均与本研究结果一致。有研究对中国其他地区山羊品种遗传分化进行研究,Fst与本研究结果范围一致[16-17]。
3.3 母系起源分析 李祥龙等[18]发现,家养山羊中既有弯角也有旋角,因此认为捻角野山羊和旋角野山羊都是家养山羊的野生祖先。实际上由于捻角野山羊的地理分布与山羊最初家养的地方有高度的相关性,所以捻角野山羊是最可能的候选者[19]。本研究发现槐山羊、济宁青山羊、伏牛白山羊、太行黑山羊和尧山白山羊均与捻角野山羊聚为一类,旋角野山羊单独聚类。本研究结果与其他中国家养山羊母系起源进化研究结果一致[15,20],均认为捻角野山羊是中国家养山羊的野生祖先。
中国山羊品种存在A、B、C 和D 4 个分支[21],A和B 是在我国山羊品种中广泛存在的类型,且经历过群体扩张,而类型C 和D 则以低频率存在于我国部分山羊个体中[15,22],本研究印证了这些观点。本研究聚类发现,中原地区山羊共分为两大支系,支系A 和支系B,且槐山羊、济宁青山羊、伏牛白山羊和尧山白山羊在A、B 支系均有分布。而太行黑山羊仅来源于支系A。当然,要澄清山羊的起源进化问题,可能仅有mtDNA D-loop标记是不够的,必须借助其他遗传标记研究。
4 结 论
中原地区5 个山羊品种线粒体D-loop 区单倍型多样度与核苷酸多样度较高,品种间均未达到亚种间遗传距离,伏牛白山羊、济宁青山羊和太行黑山羊近期经历瓶颈效应,槐山羊和尧山白山羊符合动态平衡。系统进化树结果表明,中原地区山羊祖先为捻角野山羊,有支系A 和支系B 2 个母系起源。
