泛素化在心肌梗死后的心肌保护与促再生作用的研究进展
2021-09-13刘镏王昊王连生
刘镏 王昊 王连生
(南京医科大学第一附属医院心内科,江苏 南京 210000)
目前心血管疾病仍是中国导致死亡的头号原因,其中危害最大的一种是心肌梗死[1],尽管已有包括溶栓、冠状动脉旁路移植术和经皮冠状动脉介入治疗等在内的多种治疗方法,但缺血再灌注损伤以及梗死区域心肌细胞不可逆死亡,仍无有效的治疗方法。最近的研究显示,蛋白质翻译后修饰,在心肌细胞的保护和内源性再生方面起着积极的作用[2],预示着对蛋白质修饰的干预可能是心肌梗死一个有前景的治疗手段。
泛素化是一种作用范围广泛的蛋白质翻译后修饰,在既往的研究中,泛素化更多地参与了促进凋亡、自噬的病理过程[3],但最近的研究显示,泛素化同样参与心肌梗死后的心肌细胞保护过程,如其对受损细胞器和异常蛋白质的自噬清除对心肌细胞起着保护作用[4]。同时,随心肌细胞内源性再生研究的进一步深入,以往心肌细胞不可再生的固有认知被打破[5],泛素化在这个过程中的潜力也受到越来越多的关注。现聚焦于泛素化的双重保护机制,为心肌梗死后治疗的发展提供新的见解和思路。
1 泛素与泛素化
泛素是一种低分子量蛋白质(分子量约为8.5×103),含有76个氨基酸残基,它高度保守,如酵母和人的泛素之间仅有3个氨基酸的差别[6],保证了从实验动物到人的研究结论更有普适性。泛素的功能依赖各种酶的协同,它和蛋白酶体组成泛素-蛋白酶体系统,在细胞质参与自噬和蛋白酶体介导的蛋白降解等过程[7],在细胞核参与转录调节和DNA修复等过程[8]。
经典的泛素化途径是指泛素在一系列酶的催化下和底物结合的过程,这些酶主要分为三类:E1泛素激活酶、E2泛素结合酶和E3泛素连接酶。首先E1激活泛素,然后E2将泛素转移到E3,最后E3催化泛素与靶蛋白的共价结合,其中E3决定泛素和底物连接的特异性和精确性,人类基因组里有超过800个E3[9],因此泛素化比单一基团变化的蛋白质修饰更加复杂,功能更加多样。一些和泛素具有高度相似的一级结构的蛋白质,被称为泛素样蛋白,如小分子泛素相关修饰物蛋白(SUMO)[10],它们有着与泛素相似的特性与作用过程,一样可通过修饰靶蛋白来调节靶蛋白的活性,参与许多关键的细胞过程。
2 泛素化与缺血再灌注中的心肌保护作用
2.1 外源泛素
泛素本身就可产生保护作用,一项2018年的研究证实,除抑制炎症外,外源泛素还能抑制β-肾上腺素能受体所导致的心肌细胞凋亡和纤维化,再灌注损伤3 d后观察发现心脏功能和梗死面积得到明显的改善[11]。进一步的研究发现,C-X-C型趋化因子受体4(CXCR4)是泛素的外源受体,可接受泛素的直接调节,在促进心肌细胞对氧化应激产生耐受的同时保持线粒体膜电位的稳定,减少心肌细胞的凋亡[12]。
2.2 E2泛素结合酶
尽管E2泛素结合酶在人类基因组内数量不多,但也发挥着积极的保护作用。泛素结合酶E2S可修饰β-连环蛋白,使其稳定性上升产生积累,从而促进缺氧诱导因子1α(HIF-1α)的表达,增强心肌细胞对缺血缺氧的耐受,减少心肌细胞的凋亡[13]。
2.3 E3泛素连接酶
E3泛素连接酶在人类基因组内数量庞大,其中许多E3对心肌细胞起着保护作用。Parkin是一种参与有丝分裂的E3,很早就被证实在心脏中发挥了保护作用[14],随着进一步的研究,Parkin被发现可促进亲环素D(CypD)的泛素化来下调它的表达,从而减少H2O2诱导的线粒体通透性转换孔(mPTP)的开放,稳定心肌细胞的内环境,抑制氧化应激产生的损伤,改善心功能[15]。鼠双微体2(MDM2)是一种参与调节细胞生长和凋亡的E3,据推测可通过促肿瘤抑制因子p53的泛素化降解,产生抑制心肌细胞凋亡和肥大的双重作用,这也提示抑制MDM2的抗肿瘤药物可能有不良的心脏效应[16]。类似的,泛素连接酶组成型光形态建成蛋白1(COP1)可在长链非编码RNA Fendrr的促进下,泛素化并降解细胞凋亡因子p53来减少心肌细胞的凋亡[17]。c-Jun氨基端激酶(JNK)在缺血再灌注损伤后被激活,促进心肌细胞的凋亡,c-Jun是其近端效应子,肌肉环状指蛋白1(MuRF1)是在一种作用于磷酸化激活的c-Jun并促其被26S蛋白酶体降解的E3,从而阻断损伤通路,减少心肌细胞的凋亡[18]。还有一种E3可使JNK去磷酸化,在小分子核糖核酸(miRNA)378a-3p的作用下,E3泛素连接酶三部分基序蛋白55(TRIM55)可促使双特异性磷酸酶1(DUSP1)使JNK脱磷酸化,阻断损伤通路,减少心肌细胞的凋亡[19]。损伤发生后,DNA修复酶聚腺苷二磷酸核糖聚合酶1(PARP1)会过度激活使腺苷三磷酸耗竭从而使心肌细胞凋亡,Cullin4A是E3泛素连接酶CRTs的核心,可泛素化PARP1来降低其表达,从而阻断损伤通路[20]。除上述机制外,E3泛素连接酶还会参与到已知的心肌保护通路中,如刻痕受体1(Notch1)介导的心脏保护作用,会在miRNA322对泛素连接酶F-box和WD重复结构域7(FBXW7)的抑制下得到活化,减少心肌细胞的凋亡[21]。另一个具有心肌保护作用的磷脂酰肌醇3-激酶/蛋白激酶B(PI3K-Akt)通路,会被泛素连接酶神经前体细胞表达发育下调蛋白4-1(NEDD4-1)激活,从而减少心肌细胞的凋亡[22]。
2.4 蛋白酶体
完整的泛素化过程,还包括泛素化后蛋白酶体的效应。蛋白酶体的功能不全是一个损伤因素,而增强它的功能可起到心肌保护作用[23]。可溶性晚期糖基化终产物受体(sRAGE)是一种参与多种疾病过程的内源性物质,本身就可通过信号转导及转录激活蛋白3(STAT3)途径保护心肌细胞[24],同时它也可提高蛋白酶体的活性,促进其降解异常蛋白质[25]。最近的研究也表明,泛素化和蛋白酶体的偶联也影响着心肌的保护作用,泛素1是一种蛋白酶体外泛素受体,它的上调会加强两个过程的偶联,提高降解异常蛋白的能力,改善心脏功能[26]。
2.5 泛素样蛋白
泛素样蛋白也发挥着心肌保护作用。F位点邻近转录物10(FAT10)是一种泛素样蛋白,可通过抑制凋亡因子p53及其底物miRNA34a来抑制心肌细胞的凋亡[27],FAT10被认为还可与泛素竞争结合一种具有心肌保护作用的小窝蛋白3(Cav-3)上的位点,从而减少其降解,增强心肌细胞对缺血缺氧的耐受[28]。SUMO在哺乳动物中共有4种,其中SUMO1和SUMO2在心肌细胞中起着积极的作用。锌可促进SUMO1修饰动力相关蛋白1(Drp1)[29],同时SUMO1还可在Luteolin的促进下,修饰并上调心肌钙循环的关键酶肌浆网钙三磷酸腺苷酶(SERCA2a)的表达与活性,这些都可增强心肌细胞对缺血缺氧的耐受能力[30]。激活的STAT蛋白抑制剂1(PIAS1)是一种SUMO连接酶,可通过修饰过氧化物酶体增殖物激活受体γ(PPARγ)来抑制核因子κB,减少心肌细胞的凋亡[31]。
还有其他一些泛素连接酶具有心肌保护作用,Irisin是一种可通过多种途径发挥心肌保护作用的FNDC5蛋白质裂解产物,其中一种是通过线粒体泛素连接酶(MITOL)/MARCH5通路来上调位于线粒体和内质网连接部的MITOL的活性,从而提高心肌细胞对缺氧和活性氧的耐受,减少损伤引起的细胞凋亡[32],表1总结了泛素化在缺血再灌注损伤过程中发挥的保护作用。

表1 泛素化与缺血再灌注中的心肌保护作用
3 泛素化与内源性心肌细胞再生
自从证实新生小鼠在生后7 d内心脏有完全再生能力后[33],促进心肌细胞内源性再生也成为一个可能的心肌梗死治疗方法。根据本实验室既往的相关研究显示,蛋白质翻译后修饰在再生中发挥着积极作用[34]。泛素化比一般修饰作用更加广泛,在促进内源性再生中有着更大的潜力。
3.1 细胞周期的重新开启
成熟心肌细胞再生能力的丢失很大程度上是因细胞周期的关闭,泛素化可激活细胞周期蛋白和细胞周期蛋白依赖性激酶(CDK)从而重新打开细胞周期。S期激酶相关蛋白2(Skp2)是E3泛素连接酶Skp1-Cullin-F-box的一部分,可降解抑制细胞周期蛋白的p27,促进心肌细胞再生[35],同时Skp2可直接增强核定位信号标记的细胞周期蛋白D1(D1NLS)和CDK4的效应,进一步提高再生能力[36]。p21与p27一样,可抑制CDK,Sirt1是一种组蛋白去乙酰化酶,它可使p21的赖氨酸残基脱乙酰化并激活其泛素化降解,从而促进心肌细胞再次进入细胞周期[37]。E3泛素连接酶NEDD4L可在环状RNA Nfix表达下降的情况下减少Y-box结合蛋白-1(YB-1)的泛素化,从而减少对细胞周期蛋白A2和B1的抑制,重新进入细胞周期[38]。
3.2 参与已知的再生通路
泛素化还可激活内源性再生相关的信号通路。如E3泛素连接酶Cullin7是泛素样蛋白NEDD8的底物,Cullin7可作用于Mst1/YAP通路,最终导致YAP激活,从而促进心肌细胞增殖[39],另一种E3泛素连接酶β-转导重复相容蛋白(β-TrCP)也参与对YAP的调节,可在miRNA199a-3p的抑制下减少对YAP的泛素化,激活YAP信号[40]。SUMO同样参与内源性心肌细胞的再生过程,TGF-β-Smad信号通路是一种调节细胞分化、增殖和凋亡的通路,被SUMO2修饰后抑制,可减少细胞周期的退出和抑制凋亡,从多方面促进心肌细胞的再生[41]。
3.3 具有双重效应的泛素化过程
有的泛素化相关因子参与心肌保护和内源性再生两个过程。Smad泛素化调节因子2(Smurf2)是一种E3泛素连接酶,miRNA322/503抑制其表达,从而减少Smurf2对zeste同系物增强子2(EZH2)的泛素化,以此来激活Akt/GSK3β通路,产生促进心肌细胞增殖和抑制凋亡两方面的作用[42],表2总结了泛素化在内源性心肌细胞再生过程中发挥的作用。

表2 泛素化与内源性心肌细胞再生
4 讨论与展望
以上主要是从参与泛素化过程的各种因子的角度来讨论,也可从泛素化产生的效应来讨论,图1总结了这些泛素化的效应。
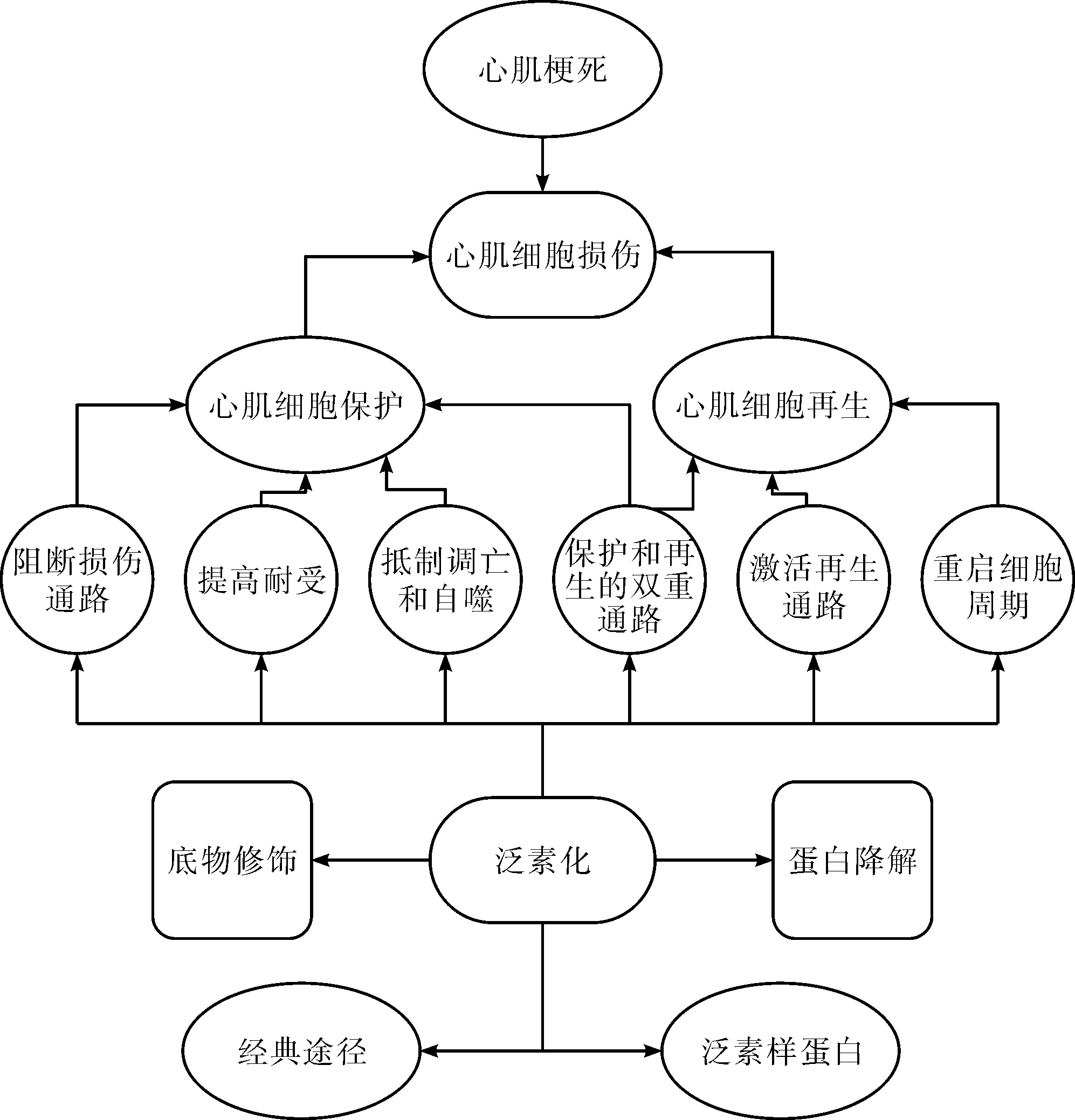
图1 泛素化参与心肌细胞保护和再生的作用效应的模式图
值得注意的是,泛素化在发挥积极作用的同时也起着介导损伤的作用,如何从众多干预靶点中找出值得进行临床转化的高效靶点,是一个仍需探索的问题。笔者也发现,同样的泛素化过程可能在发挥保护作用时也发挥促再生的作用,所以在探索促再生途径时,已有的心肌保护途径值得再次探究。总的说来,目前泛素化的研究还局限在动物模型中,还需进一步的探索来实现临床转化。
利益冲突所有作者均声明不存在利益冲突。