贵州中部避雨栽培对甜樱桃雌雄配子体形态发育的影响*
2021-08-29吴亚维冯建文韩秀梅
宋 莎,吴亚维,冯建文,韩秀梅,赵 凯
(贵州省农业科学院果树科学研究所,贵阳 550006)
甜樱桃(Prunus aviumL.)又名欧洲甜樱桃、大樱桃,果实个大、硬度高,近些年浙江、江苏、上海、湖北等地均在引种,尤其是与贵州省相邻的云南和四川甜樱桃产业的发展,使贵州省很多企业和果农受到影响也开始尝试种植。但“花而不实”、坐果率低和落果严重等问题限制了贵州省甜樱桃产业的发展。
前些年,需冷量不足通常被认为是温暖地区甜樱桃产业发展迟滞的主要限制因素[1]。近些年发现,国内比较温暖的南方地区每年冬季0~7.2 ℃的低温累积量可以使中需冷量品种顺利通过休眠[2],冬季低温不足并不是南方地区甜樱桃坐果率低的关键因素,而春季开花坐果期的降雨会直接影响甜樱桃授粉和坐果[3],夏季雨水偏多会影响花芽分化数量和质量[4]。贵州省科研工作者自2008 年起对省内引进的甜樱桃开展观察和试验研究,发现影响贵州省甜樱桃开花坐果的主要因素是降水较多,且自2015 年起在贵阳市开展了甜樱桃避雨栽培试验,已报道避雨栽培虽然影响甜樱桃的光合速率[5],但有利于花束状果枝和花芽的形成[6],也可提高植株开花数量和花器官质量,现需进一步研究避雨栽培对其雌配子体发育的影响。果树雌雄配子体正常发育是授粉受精成功的基础,在蔷薇科果树中,雌配子体发育异常会导致坐果率低[7],因而研究果树雌配子体的发育具有重要的实践意义。
果树雌雄配子体发育的相关研究较多,桃[8]、番荔枝[9]、枣[10-11]等均有报道。朱东姿等[12]应用激光共聚焦扫描显微镜观察甜樱桃‘美早’和‘布鲁克斯’的雌配子体发育过程,并统计了胚囊发育情况,推测‘美早’胚囊容易败育而‘布鲁克斯’胚囊活力高。张雪等[13]利用常规石蜡切片法观察‘马哈利’樱桃的雌雄蕊发育过程,探讨了其授粉结实率低的解剖学原因。王玉等[14]统计了甜樱桃‘拉宾斯’‘艳阳’和‘马哈利CDR-1’自花授粉坐果率,并通过石蜡切片法对其雌雄配子体发育过程进行了观察。
本试验以避雨栽培的‘布鲁克斯’和‘萨米脱’2个甜樱桃品种为试材,以露地栽培为对照,通过石蜡切片方法观察了甜樱桃雌配子体形态发育过程,分析了避雨栽培对甜樱桃雌雄配子体发育的影响,以期为探讨贵州省甜樱桃露地栽培坐果率低的原因提供科学依据。
1 材料与方法
1.1 试验材料
试验地位于贵州省农业科学院果树科学研究所贵阳市乌当区百宜镇果树试验基地,海拔1 330 m,年平均气温14.30 ℃,平均年降水量1 240 mm,年日照时数1 300~1 400 h。避雨设施为四连栋大棚,顶膜两侧可卷起1.5 m,侧膜可卷至距离地面2.0 m 高处。供试品种为5 年生‘布鲁克斯’和‘萨米脱’,砧木为‘吉塞拉6号’,行株距4.0 m×2 m,树形采用纺锤形[7]。避雨栽培和露地栽培的‘布鲁克斯’和‘萨米脱’2个甜樱桃品种各随机选取5 株树挂牌试验。
1.2 试验方法
2019—2020 年,自2 月下旬花芽开始萌动起至3 月下旬,每3 d 采1 次花芽(后为花蕾),每次随机在每株挂牌树上采10个发育良好、无病虫害的花芽或花蕾,FAA 固定液固定,常规石蜡切片法制片,番红固绿染色,NIKON ECLIPSE E100 型正置光学显微镜观察并拍照,记录形态特征和配子体发育时期。观察胚囊形态,统计胚珠和胚囊发育正常、有胚珠无胚囊、无胚珠无胚囊的花朵数量,计算百分比。
2 结果与分析
2.1 避雨栽培对甜樱桃花蕾形态发育过程的影响
通过观察不同栽培模式下甜樱桃花蕾纵切图(图1)发现,避雨栽培模式下‘布鲁克斯’和‘萨米脱’的雌配子体小孢子母细胞3 月13 日开始发育,雄配子体3 月7 日开始发育;露地栽培模式下甜樱桃雌配子体小孢子母细胞3 月16 日开始发育,雄配子体3 月10 日开始发育。对2种栽培模式下的完整花蕾切片各选了100个进行观察发现,避雨栽培模式下甜樱桃花蕾形态发育基本正常,露地栽培模式下7%的甜樱桃花蕾切开后内部组织细胞变形(图1-K)或降解(图1-L)。可见,避雨栽培可使甜樱桃雌雄配子体发育时期提前,并促进其正常形态发育。

图1 避雨栽培模式下‘萨米脱’花蕾纵切形态
2.2 避雨栽培对甜樱桃雌配子体形态发育的影响
利用石蜡切片法未能观察到甜樱桃雌配子体形态发育全过程,仅能看到3 月13—16 日子房开始发育。部分可以观察到胚珠和胚囊的存在以及形态是否正常,但很难在一张切片中同时观察到胚囊发育过程中的几个核,无法界定具体时期。对2种栽培模式下的完整子房切片各选了100个进行观察,发现雌配子体主要为正常胚珠和胚囊(图2-A)、无胚囊有胚珠(图2-B)、无胚珠无胚囊(图2-C)3种形态。根据统计结果(表1),避雨栽培模式下,‘布鲁克斯’和‘萨米脱’的正常发育胚囊均在50%以上;露地栽培模式下,‘布鲁克斯’和‘萨米脱’的正常发育胚囊均小于15%。可见,避雨栽培可提高甜樱桃正常胚珠、胚囊的比例。

图2 ‘萨米脱’雌配子体的3种形态

表1 避雨栽培对甜樱桃胚珠、胚囊形态的影响
2.3 避雨栽培‘萨米脱’雄配子体形态发育过程
由避雨栽培‘萨米脱’的雄配子体切片(图3)可以看出,甜樱桃雄配子体发育过程主要为造孢细胞时期、小孢子母细胞时期、小孢子母细胞减数分裂时期、小孢子单核居中时期、小孢子单核靠边时期、二核花粉时期、成熟花粉时期等。造孢细胞孢原细胞是上年分化形成的,2 月26 日处于造孢细胞时期,造孢细胞体积较大,细胞核大,呈不规则多边形,其他细胞没有明显变化(图3-A);3 月7日进入小孢子母细胞时期,细胞体积增大,细胞核更明显,细胞的棱角钝化变圆,已经形成小孢子母细胞,花药壁绒毡层细胞大多处于有丝分裂期,体积明显增大(图3-B);3 月10 日小孢子母细胞已经进入减数分裂时期,能看到四分体排列为四面体形(图3-C);3 月13 日四分体已分离成单个小孢子,小孢子有1个核位于中央,液泡不明显,细胞质浓(图3-D);3 月16 日小孢子体积增大,细胞内形成中央大液泡,细胞核在细胞一侧,部分细胞核形成了2个核即生殖核和营养核,进入二核阶段(图3-E),这一时期进行时间较长;到3 月25日开始形成成熟的二核花粉。
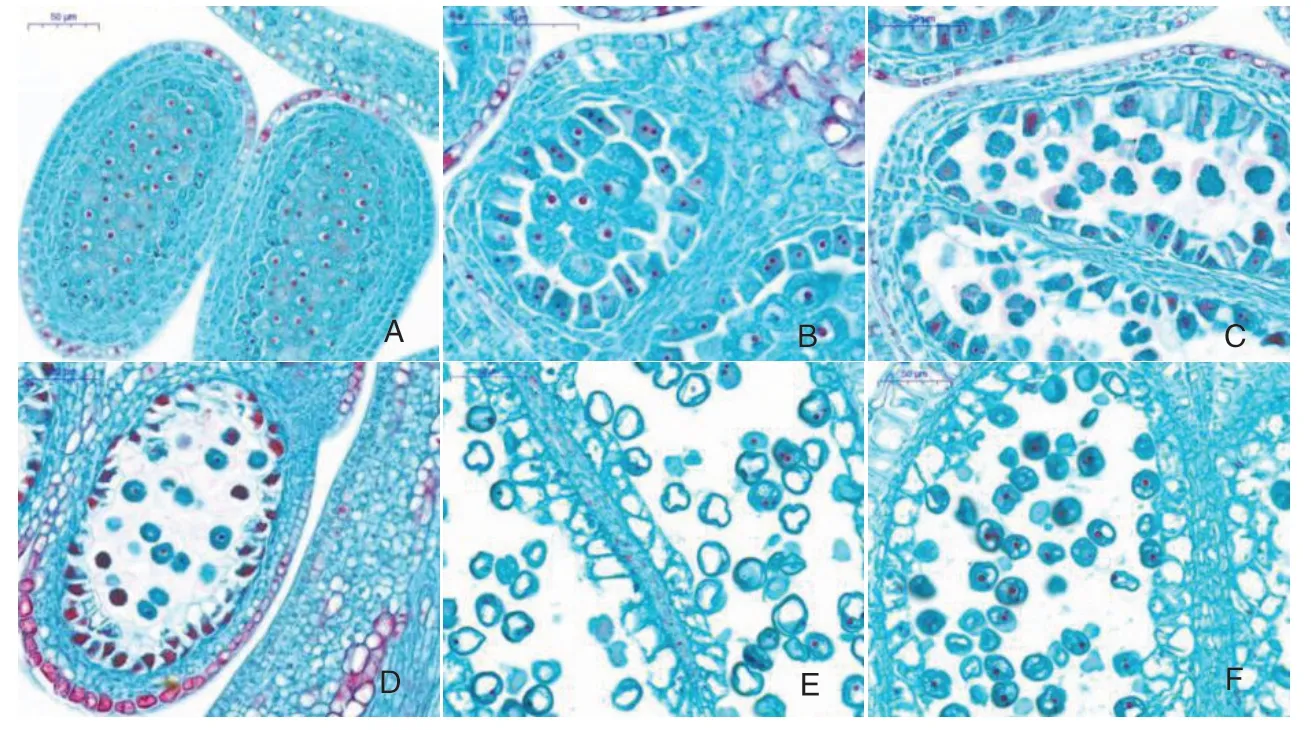
图3 避雨栽培模式下‘萨米脱’雄配子体形态发育过程
2.4 避雨栽培对甜樱桃雄配子体形态发育的影响
同一日期采集的不同栽培模式、不同品种甜樱桃的花蕾雄配子体发育时期不同。由表2 可知,避雨栽培模式下,‘布鲁克斯’和‘萨米脱’的雄配子体发育时期在3 月13 日之前基本相同,之后‘萨米脱’稍早于‘布鲁克斯’进入小孢子单核靠边期;露地栽培模式下,‘布鲁克斯’进入小孢子母细胞、小孢子母细胞减数分裂、小孢子单核居中等时期均稍早于‘萨米脱’,而形成成熟花粉的时期基本一致。避雨栽培模式下2个甜樱桃品种的雄配子体的几个发育时期均早于露地栽培,但到3 月28 日均形成了成熟花粉。对2种栽培模式下3 月28 日的完整花药切片各选了100个进行观察发现,露地栽培模式下甜樱桃雄配子体会发生进入小孢子母细胞时期后不再进行减数分裂(图4-A)、逐渐降解(图4-B)或小孢子时期形成空花粉囊(图4-C)等现象使雄配子体败育,其中小孢子母细胞不进行减数分裂或逐渐降解的占5%,空花粉囊占4%;而避雨栽培模式下雄配子体形态发育正常。可见,避雨栽培不仅促使甜樱桃雄配子体发育时期提前,还能促进雄配子体正常发育。

表2 避雨栽培对甜樱桃雄配子体形态发育时期的影响
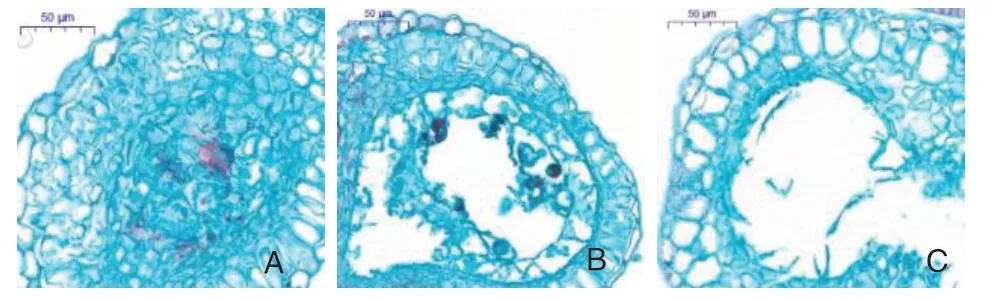
图4 露地栽培模式下‘萨米脱’雄配子体败育形态
3 讨论与结论
植物雌配子体发育与受精存在直接关系,雌配子体发育不良将导致不能正常受精与坐果[15]。胚囊是雌性器官的重要组成部分,也是受精和胚珠形成种子的重要场所,胚囊异常发育会导致无法正常受精发育成果实。朱东姿等[12]应用激光共聚焦扫描显微镜清楚地观察了甜樱桃雌配子体的大孢子母细胞时期、四分体时期、一核胚囊时期、二核胚囊时期、四核胚囊时期、八核胚囊时期等6个发育时期,发现‘布鲁克斯’和‘美早’的胚囊败育率分别为21%和41%。因采样时间和石蜡切片方法的限制,未能在同一张切片中同时观察到胚囊发育过程中的几个核,无法界定甜樱桃雌配子体具体发育时期,本研究中未能很好地观察到雌配子体的发育过程,仅根据观察到的情况统计了雌配子体胚珠和胚囊的存在情况。避雨栽培模式下甜樱桃形态发育正常的胚囊、胚珠在50%以上,而露地栽培模式下小于15%;雌配子体发育成有胚珠无胚囊类型比例较高,是造成雌配子体败育的重要原因。本研究中甜樱桃雄配子体的形态发育过程主要有造孢细胞时期、小孢子母细胞时期、小孢子母细胞减数分裂时期、小孢子单核居中时期、小孢子单核靠边时期、二核花粉时期、成熟花粉时期等,这与王玉等[14]的观察结果基本一致,但雄配子体发育过程中有9%左右出现未进行减数分裂、降解或空花粉囊现象。雌配子体胚囊败育和雄配子体发育过程中降解可能是露地栽培甜樱桃落花落果的主要原因。
花器官是与甜樱桃产量关系最密切的器官,它的正常发育是获得高产的关键[16]。开花期统计露地栽培‘布鲁克斯’和‘萨米脱’开花数量,每株仅60 朵左右;实际上,冬季统计露地栽培甜樱桃结果枝和花芽数量时并不仅仅形成几十个花芽[6]。这与试验地气候条件有关。自2 月26 日至3 月9 日,试验地露地日平均气温高于避雨设施内,甜樱桃雌雄配子体发育较慢;3 月10 日起,避雨设施内日平均气温高于露地,小孢子母细胞进入减数分裂时期开始快速发育;3 月25—28 日是日平均气温较高的几天,雄配子体形态发育基本完成,形成成熟的二核花粉。可见,在甜樱桃雌雄配子体发育过程中低温可能会导致发育不完全或畸形现象出现。试验地冬季降雨或大雾天气较多,到翌年2 月花芽开始萌动前,因生长环境的湿度持续较大对露地栽培甜樱桃花芽发育产生了不可逆伤害,部分花芽腐烂或停止生长,从而发生本研究中外部正常的花蕾切开后内部结构均已降解的现象。自2 月26 日至3 月28日,仅有3~4 d 避雨设施内湿度略高于露地,其余时间避雨设施内湿度比露地低;避雨设施内虽然湿度降低幅度不大,但甜樱桃发育的花蕾上没有水滴,大多处在干燥状态,大多可正常发育;而露地栽培甜樱桃的花芽、花蕾等在湿度较大时整个组织处于潮湿状态,进行形态发育的花蕾易出现畸形现象,还有部分花芽腐烂。可见,冬春季降雨是影响贵州省中部露地栽培甜樱桃开花结实的重要因素。
虽然避雨设施影响了甜樱桃的光合速率[5],但避雨栽培明显促进甜樱桃花芽形成[6],也提高了雌雄配子体正常发育的比例,贵州省中部避雨栽培甜樱桃意义重大,降低避雨设施成本以及提高覆盖材料透光率的技术值得进一步研究。本研究虽然对甜樱桃雄配子体的形态发育状况观察得较为完善,但由于选片没能对雌配子体进行仔细的研究,对甜樱桃雌配子体形态发育过程只能提供相对可靠的数据。在今后的工作中对甜樱桃胚珠、胚囊及后续的胚胎形态发育等问题有待解决。
