铁死亡调控机制的研究进展
2021-08-24马臣杰宋孚洋
马臣杰,张 雯,曾 瑾,宋孚洋
(1. 宁夏大学 生命科学学院 西部特色生物资源保护与利用教育部重点实验室,银川 750021;2. 宁夏回族自治区兽药饲料监察所,银川 750001)
细胞死亡(Cell death)与生物体的生长、发育、衰老和疾病密切相关。迄今为止,人们已经发现凋亡(Apoptosis)、程序性坏死(Necroptosis)、焦亡(Pyroptosis)和氧中毒(Oxeiptosis)等多种程序性细胞死亡(Programmed cell death,PCD)。Dolmsy等[1]在筛选抗肿瘤药物时发现了一种能够引起癌细胞死亡的新化合物Erastin。有趣的是,Erastin诱导的细胞死亡方式与当时已知的细胞死亡方式皆不相同,未观察到凋亡小体的形成、DNA的断裂以及Caspase家族的激活,亦无法被Caspase抑制剂逆转。经过数年深入研究,Dixony等[2]于2012年进一步提出了一种铁依赖性的程序性细胞死亡概念,并将其正式并命为铁死亡(Ferroptosis)。
铁死亡的发生伴随着细胞内铁离子流紊乱、活性氧(ROS)和脂质过氧化物水平的显著性升高等现象,最终因细胞膜磷脂内侧发生脂质过氧化反应导致膜破损,细胞崩解死亡。已经证实铁死亡与神经退行性疾病、肿瘤疾病、心脑血管疾病和急性肾损伤等多种疾病的存在密切关系。
1 铁死亡的特征
1.1 铁死亡的细胞形态学特征
铁死亡形态学特征主要包括:线粒体膜密度增高、脊显著减少、膜出现破裂,整体表现为线粒体萎缩;细胞核大小正常且保持完整、染色质未见发生凝集;细胞膜未见起泡,但膜密度增加;膜磷脂双分子层结构和细胞膜的流动性均发生了改变[3](表1)。
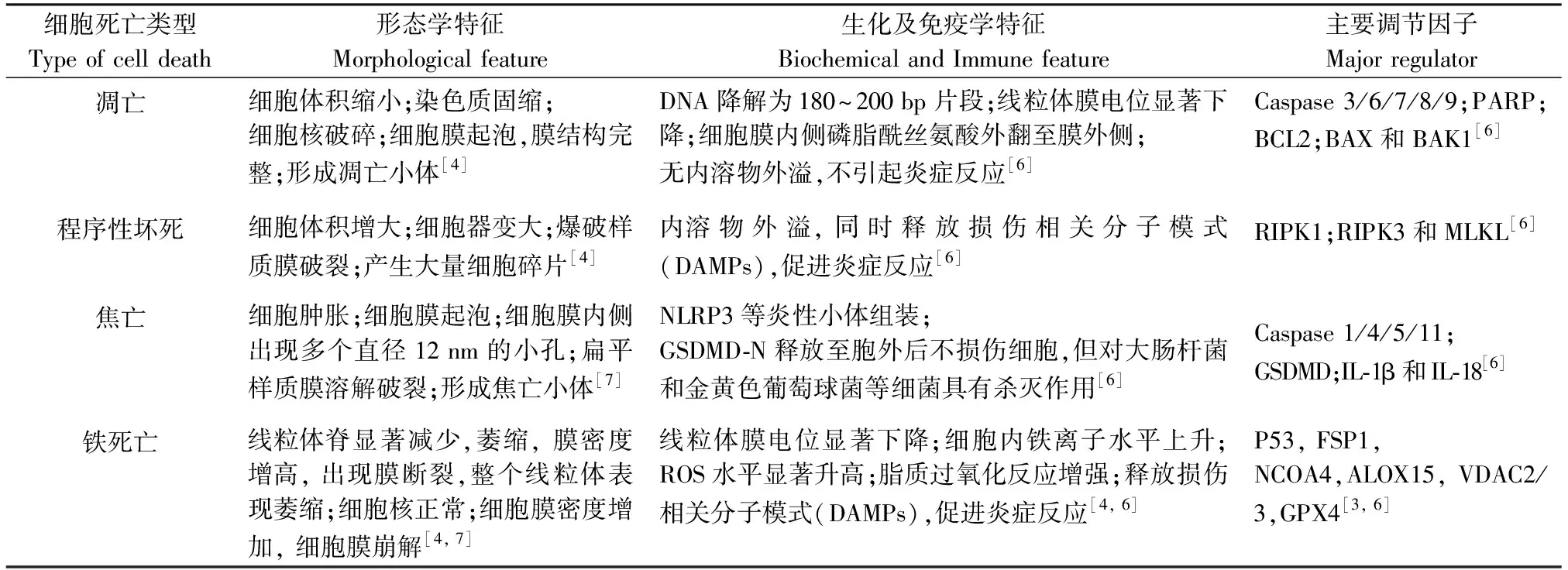
表1 铁死亡与凋亡、程序性坏死及焦亡之间的特征比较Table 1 Characteristic comparison among ferroptosis,apoptosis,necroptosis and pyroptosis
1.2 铁死亡的生物化学特征
铁死亡所具有典型的生化特征,常被用来评价铁死亡的发生状态。如:细胞内Fe2+和活性氧(ROS)水平的升高发生芬顿反应(Fenton reaction),半胱氨酸摄入量的减少,谷胱甘肽的耗竭,羟基(OH-)、羟基自由基(·HO)和脂质氢过氧化物(Lipid hydroperoxides,LOOH)的水平升高,生物膜中多聚不饱和脂肪酸发生脂质过氧化反应等[4]。
1.3 铁死亡的调控因子
研究发现,参与调节铁死亡的关键细胞因子大致有数十种[5],主要包括核受体辅激活蛋白4(Nuclear receptor coactivator 4,NCOA4)、花生四烯酸15-脂氧合酶(Arachidonate 15-Lipoxygenase,ALOX15)、Voltage-dependent anion-selective channel protein 2(VDAC2/3)、SLC7A11、SLC3A2、谷胱甘肽过氧化物酶4(Glutathione peroxidase,GPX4)、转铁蛋白受体(Transferrin receptor,TFR)、转铁蛋白(Transferrin TF)及储铁蛋白(Ferrtin)等[6]。
2 铁死亡的分子调控机制
2.1 铁离子代谢
铁元素是生物体必需的营养元素之一。通常细胞内铁的平衡性表现在铁的吸收、输出、利用和储存之间的平衡。当铁死亡发生时,细胞内积累大量的游离Fe2+。游离Fe2+氧化性极强,容易与H2O2发生芬顿反应(Fenton reaction),产生能对DNA、蛋白质和膜脂质造成氧化损伤的羟基自由基,促进脂质过氧化反应的发生损伤细胞膜导致细胞死亡[8]。芬顿反应式为Fe2++H2O2=Fe3++OH-+·HO。
已知铁死亡发生时Fe2+的主要来源有2个:(1)携带Fe3+的转铁蛋白(Transferrin, TF)与质膜上的转铁蛋白受体(Transferrin receptor,TFR)结合后,质膜形成囊泡将携带Fe3+的转铁蛋白摄入胞内,随后囊泡内低pH促使Fe3+从转铁蛋白剥离,脱落的Fe3+被还原成Fe2+游离于胞质中,或者与Ferritin结合形成铁池(Iron pool);(2)铁池中的铁蛋白可在核受体辅激活蛋白4(Nuclear receptor coactivator 4,NCOA4)的介导下被自噬溶酶体包裹,进而降解、释放出大量Fe2+(图1)。
2.2 脂质过氧化反应
脂质过氧化(Lipid peroxidation)是指脂质在自由基或脂质过氧化酶的作用下失去的氢原子,导致脂质碳链的氧化、断裂及缩短,并产生脂质自由基、脂质氢过氧化物和活性醛(丙二醛、4-羟基壬烯醛)等具有细胞毒性物质,最终造成细胞损伤的脂质氧化降解反应[9]。在铁死亡中,脂质过氧化的危害主要体现在它导致了多聚不饱和脂肪酸(Polyunsaturated fatty acid,PUFA)和磷脂酰乙醇胺(Phosphatidylethanolamines PE)2种重要生物膜组分的氧化降解。
PUFA是细胞和细胞器膜磷脂的主要成分,也是合成磷脂双分子层内层主要成分PE的重要底物。磷脂双分子层作为维持细胞膜流动性的结构基础,对维持细胞的正常生长、增殖、分化、衰老、死亡、免疫等具有重要功能。脂质过氧化反应会改变PUFA的分子构型,破坏细胞膜结构的流动性和稳定性,细胞膜通透性增加,细胞容易破裂死亡。PUFA与自由基亲和力高,其双键之间的氢原子极易被自由基氧化。PUFA的脂质过氧化反应大致分为2个阶段。首先,羟基自由基和过氧化氢等活性氧类物质获取PUFA中的氢原子产生脂质自由基(Lipid ROS,L-);然后,脂质自由基与氧分子作用生成脂质过氧化自由基(Lipid peroxyl radicals,LOO-)。脂质过氧化自由基从其他PUFA中夺取氢原子形成脂质自由基和脂氢过氧化物(Lipid hydroperoxide,LOOH)。脂质过氧化自由基能不停参与PUFAs的氧化过程,使得PUFAs的脂质过氧化反应具有级联式反应特征。
PE是生物膜甘油磷脂,在线粒体内膜中含量约占总磷脂的40%,在其他细胞器生物膜中约占总磷脂的15%~25%[10]。PE具有多种细胞功能,包括作为磷脂酰胆碱的前体和重要的翻译后修饰的底物,影响膜拓扑结构,促进细胞和细胞器膜融合、氧化磷酸化、线粒体生物生成和自噬等[11]。PE与自由基的亲和力并不高,在发生脂质过氧化之前需要在2种酶作用下形成氧化位点。首先,酰基辅酶a合成酶长链家族成员4(Long-chain acyl-CoA synthetase-4,ACSL4)利用花生四烯酸(Arachidonic acid,AA)和肾上腺酸(adrenic acid,AdA)合成花生四烯酰-CoA(AA-CoA)和肾上腺酰-CoA(AdA-CoA);然后,AA/AdA-CoA在溶磷脂酰胆碱酰基转移酶3(Lysophosphatidylcholine Acyltransferase 3,LPCAT3)催化下与PE形成PE-AA/AdA[12]。PE-AA/AdA容易被自由基或花生四烯酸15-脂氧合酶(Arachidonate 15-Lipoxygenase,ALOX15)氧化为具有细胞毒性的脂氢过氧化物PE-AA/AdA-OOH促进铁死亡发生[12](图1)。
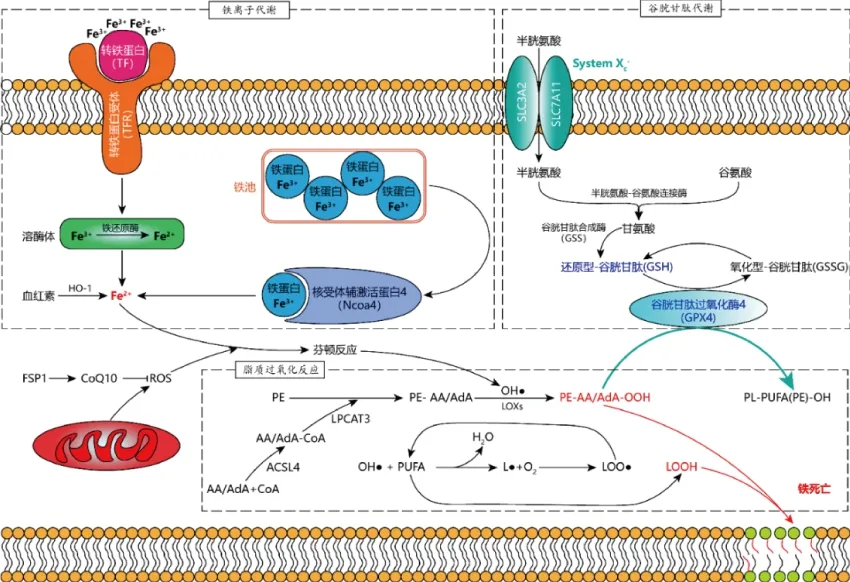
图1 铁死亡的分子调控机制Figure 1 Mechanisms of ferroptosis
值得一提的是,Doll等[13]和Bersuker等[14]发现线粒体凋亡诱导因子2(Apoptosis inducing factor mitochondria-associated 2,AIFM2)能与脂质过氧化自由基清除剂——辅酶Q10(Coenzyme Q10,CoQ10)协同作用抑制细胞铁死亡的作用,并将其更名为铁死亡抑制蛋白1(Ferroptosis-suppressor-protein 1,FSP1)。
2.3 谷胱甘肽代谢
谷胱甘肽是一种水溶性三肽,由谷氨酸、半胱氨酸和甘氨酸氨基酸残基组成。人体中的谷胱甘肽有2种型态,一种是还原型谷胱甘肽(GSH),另一种是氧化型谷胱甘肽(GSSG)。还原性谷胱甘肽是人机体内重要的抗氧化剂,不仅将H2O2还原成H2O,清除自由基,维持胞内自由基含量的平衡状态,还作为谷胱甘肽过氧化物酶4(Glutathione peroxidase,GPX4)的辅因子参与过氧化脂质LOOH的还原反应,修复生物膜中的LOOH,防止铁死亡的发生。谷胱甘肽的耗竭会导致GPX4失活,增加细胞内脂质过氧化作用,从而导致铁死亡的发生。GPX4是细胞内最重要的抗脂质过氧化酶,亦是铁死亡的核心调控因子。GSH作为GPX4的辅因子,可与GPX4协同发挥抗脂质过氧化反应的作用,促使细胞膜上的脂质过氧化物发生还原反应,遏制铁死亡的发生。因此,谷胱甘肽耗竭,GPX4酶活被抑制,是细胞执行铁死亡的必要条件。
细胞主要通过合成谷胱甘肽来维持胞内谷胱甘肽水平。谷胱甘肽的合成过程主要分为2步(图1):第1步,细胞按照1∶1的摄入比例通过System Xc-从胞外摄入半胱氨酸和谷氨酸,半胱氨酸和谷氨酸在依赖于ATP的半胱氨酸-谷氨酸连接酶(Gglutamate-cysteine ligase,GCL)作用下形成γ-谷氨酰半胱氨酸;第2步,γ-谷氨酰半胱氨酸和甘氨酸经谷胱甘肽合成酶(Glutathione synthetase,GSS)催化生成谷胱甘肽。谷胱甘肽的合成效率主要受限于半胱氨酸的浓度。
溶质载体家族7成员11(SLC3A2)和溶质载体家族3成员2(SLC7A11)组成了半胱氨酸/谷氨酸转运蛋白系统(System Xc-)。当System Xc-亚基活性被抑制时,细胞对半胱氨酸的摄入量不足,进而阻断了谷胱甘肽的合成,致使被H2O2和过氧化脂质迅速消耗的谷胱甘肽无法得到补充;同时,胞内谷胱甘肽浓度下降会抑制GPX4的活性,导致细胞抗脂质过氧化能力骤降,细胞极易发生铁死亡。高明辉等[15]研究证实,细胞处于半胱氨酸饥饿状态时,清除线粒体或者抑制线粒体电子转移链(Electron transfer chain,ETC)可减弱铁死亡的发生。
3 铁死亡的调节剂
3.1 铁死亡的激活剂
依据铁死亡激活剂的作用靶点,大致可以将其分为3类。第1类是System Xc-抑制剂,例如Erastin和Sorafenib等。Erastin是铁死亡强效激活剂,一方面它通过System Xc-阻断细胞对半胱氨酸的摄入,造成谷胱甘肽(Glutathione,GSH)耗竭,降低GPX4活性,进而减弱细胞抗脂质氧化能力[1-2];另一方面它与线粒体电压依赖阴离子通道蛋白(Voltage-dependent anion-selective channel protein 2,VDAC2/3)结合并抑制其活性,导致线粒体功能障碍并释放大量的自由基,增强细胞脂质过氧化反应[16]。Sorafenib则是一种广谱激动剂,除了诱导铁死亡之外,还可引起细胞自噬和凋亡[17]。第2类是GPX4抑制剂,例如RSL3和FIN56等。RSL3可与GPX4结合并抑制其蛋白活性,导致胞内脂质过氧化物积累过量[18]。FIN56是一种特异性较强的铁死亡诱导剂,可降低胞内GPX4和CoQ10蛋白丰度来激活铁死亡[19]。RSL3和FIN56都可直接作用于抗脂质过氧化核心细胞因子GPX4。第3类是GSH消耗剂,例如半胱氨酸酶(Cysteinase)、BSO和顺铂(Cisplatin)等。Cysteinase可直接降解半胱氨酸,阻断GSH的合成[20]。BSO抑制谷氨酸半胱氨酸连接酶的活性,导致胞内无法产生足量的GSH合成前体——甘氨酸[21]。顺铂是一线广谱抗癌药物,其主要副作用之一是增加机体内ROS水平消耗GSH,具有肾毒性[22]。
此外,有些激活剂可以通过诱导脂质过氧化反应或者增加胞内游离铁离子水平来触发铁死亡。如iFSP1可在过表达FSP1的GPX4基因缺失细胞系中抑制FSP1活性并促进脂质过氧化反应对细胞造成损伤[13]。枸橼酸铁铵通过上调胞内游离铁离子水平,来强化芬顿反应产生过量的自由基[23]。
3.2 铁死亡的抑制剂
鉴于铁死亡的发生依赖铁离子的代谢紊乱,以及脂质或谷胱甘肽的过氧化反应,所以大部分铁死亡抑制剂属于铁螯合剂或抗氧化剂。铁螯合剂的主要作用是清除游离的铁离子。甲磺酸去铁胺(Deferoxamine mesylate,DFO)是临床上使用最多的铁离子螯合剂之一,可与游离铁离子结合形成稳定的复合物,从而削弱芬顿反应,常被用于治疗慢性铁过载相关疾病[24]。铁死亡抗氧化剂能够清除羟基自由基或脂质自由基。最经典的自由基清除剂为他汀类药物和利普生司他汀类药物。初代他汀类药物Ferrostatin是目前使用最广泛的铁死亡强效抑制剂,可以阻止Erastin诱导的细胞内ROS和脂质ROS的积累[2]。利普生司他汀类药物Liproxstatin含有酰胺类和磺胺类亚基,在机体内具有良好的稳定性和药物吸收分布[25]。尚未发现Liproxstatin除了抑制铁死亡之外能否影响其他细胞死亡方式[26]。
脂质过氧化酶抑制剂也具有抑制铁死亡的作用。例如脂溶性抗氧化剂维生素E(Vitamin E),不但可以抑制LOX活性,而且其O-H键较弱容易向过氧自由基和其他自由基输送氢原子,保护细胞免受铁死亡氧化损伤[27]。哮喘药齐留通(Zileuton)可抑制LOX-5水平,可通过减弱PE的脂质过氧化反应降低铁死亡的发生[28]。
4 铁死亡与疾病的关系
4.1 癌症(Cancer)
癌症是一类几乎可以发生在人体的任何器官或组织中,并且容易侵袭扩散至临近组织器官的疾病,被世界卫生组织列为人类第二大死亡病因[29],给全球患者和医疗卫生系统造成了巨大伤害和损失。促进肿瘤细胞凋亡一直是研发抗癌药物的重点方向,然而癌细胞具有较强的抗凋亡能力,使得这类抗癌药物在实际应用中疗效有所弱化。
近年来,在大肠癌[30]、胃癌[31]、胰腺癌[32]、肺癌[33]和三阴性乳腺癌[34]等癌症研究中,发现诱导肿瘤细胞发生铁死亡或者铁死亡激活剂与抗肿瘤药物联合作用时利于对肿瘤的干预、抑制和清除[30-32]。
4.2 神经退行性疾病(Neurodegenerative disease)
神经退行性疾病是一类因神经元结构或功能逐渐退化导致的疾病。神经元是大脑和脊髓的基本组成单位,它的病变会给大脑和脊髓造成不可逆的损伤,影响人体运动或记忆功能。在大脑的老化过程中,黑质部、球状核、尾状核、皮层等脑部区域会出现进行性的铁沉积,而这些大脑区域与神经退行性疾病密切相关[35]。阿尔茨海默病(AD)是最常见的认知功能障碍神经退行性疾病,其重要病理特征之一是脂质过氧化物的积累、铁离子稳态失衡[36]及海马体严重受损[37]。研究发现,缺失GPX4基因的小鼠同时具有铁死亡生化特征和AD表征,并且该小鼠模型的AD表征可以被铁死亡抑制剂缓解[37]。神经元发生病变死亡,其病变神经元中显示铁水平激增[38]和抗氧化剂GSH耗竭[39]。临床研究表明,铁螯合剂可以通过降低患者体内的铁含量来缓解早期PD患者的运动症状,从而减轻患者的运动症状[40]。
4.3 缺血再灌注损伤(Ischemia/reperfusion injury)
缺血再灌注损伤是指缺血的组织或器官经血液灌注后发生结构损伤和功能障碍的症状。铁死亡在缺血再灌注损伤中发挥了重要的调节作用[41]。目前,多种铁死亡相关治疗剂已被用于治疗缺血再灌注引起的肠道损伤[42]和横纹肌溶解症(Rhabdomyolysis)[43]等疾病。铁死亡抑制剂 Liproxstatin可改善缺血/再灌注所致小鼠肝损伤[44]。
4.4 急性肾损伤(Acute kidney injury,AKI)
急性肾损伤患者常常会伴随突然的肾脏功能丧失,通常作为急性肾损伤的并发症而发生,严重时导致患者死亡。研究发现,铁死亡抑制剂在AKI模型中具有良好的肾脏保护效果[45-46]。
5 小结
铁死亡作为一种由铁离子、自由基及脂质过氧化物介导的非凋亡性程序性细胞死亡方式,它的起始和执行都是在严格的分子调控机制下进行。尽管已有许多研究成果证实了铁死亡与多种疾病具有密切的关系,但是铁死亡在人类疾病中的功能研究尚处于起步阶段。铁死亡如何促进炎症的发展?铁死亡与免疫应答是否存在关联?仍有许多未知问题有待解答。深入研究铁死亡在人类疾病中的作用,对寻求更有效的疾病干预和治疗药物具有深远意义。
