高蛋白食品中杂环胺形成与控制的研究进展
2021-06-01陈珍妮姚利利刘德果史莉莉赵天培
席 俊,陈 阳,陈珍妮,姚利利,刘德果,史莉莉,李 潘,赵天培
河南工业大学 粮油食品学院,河南 郑州 450001
热传递是食品热加工的重要目的,可以达到杀菌、熟化、提供独特风味等效果,而高蛋白食品在热加工过程中,会因蛋白质的氧化、降解、聚集等产生许多危害物,包括晚期糖基化终末产物、丙烯酰胺、杂环胺等[1-2]。毒理学实验表明杂环胺毒性远超典型致癌物黄曲霉毒素B1、多环芳烃、亚硝酸盐等[3-5]。目前从食品基质中分离的杂环胺类物质超过30种,在食品中广泛存在,在经高温烹制的肉食制品中尤为显著[6]。国际癌症研究机构IARC将12种杂环胺归为2B组致癌物,将2-氨基-3-甲基-3H-咪唑并[4,5-f]喹啉(IQ)归为2A组致癌物。杂环胺的致突变基础机制已经得到很好的表征,细胞色素P450酶将杂环胺转化为羟胺,通过N-乙酰基转移酶或磺基转移酶再转化为相应的酯[7-8]。这些酯可以自发水解,产生与DNA共价结合的反应芳基氮离子,杂环胺-DNA加合物易于发生移码突变和点突变[9]。作者详细探讨了包括前提物在内影响杂环胺生成的主要因素,提出使用热处理植物基蛋白作为模拟体系,研究杂环胺生成与蛋白在高温下反应的相关性,来探索蛋白食品中杂环胺生成机制及对应的抑制机理的设想,以期为后续研究杂环胺的生成及抑制提供新的思路。
1 杂环胺的种类
依据生成条件将杂环胺分为两类[10-11]:氨基-咪唑-氮杂芳烃类杂环胺一般可在100~250 ℃之间形成,均有一个N-甲基-氨基咪唑主体,按结构细分为喹喔啉类、喹啉类和吡啶类,结构如表1所示;氨基咔啉类杂环胺按结构细分为α-咔啉类、β-咔啉类、γ-咔啉类、δ-咔啉类,结构如表2所示。
2 杂环胺形成的影响因素
2.1 前体物对杂环胺形成的影响
杂环胺的前体物包括氨基酸、还原糖、肌酸(酐),且各前体物在不同杂环胺的形成过程中的参与程度也不尽相同。鄢嫣[10]的试验研究表明,前体物在杂环胺生成前大量降解,肌酸的降解尤为显著,与其需脱水生成肌酐有关,作为肌酸的水解产物,肌酐的含量则是先上升后下降。早期试验发现单独热解色氨酸、谷氨酸、大豆蛋白等可产生杂环胺[12-13],后续试验发现在日常烹饪能够达到的温度下,苯丙氨酸、色氨酸和大豆蛋白等也可产生杂环胺[14-15]。肌酸(酐)通常存在于肌肉组织中,是多种杂环胺形成的前体物质,尤其是对于IQ和IQx型杂环胺来说,肌酸(酐)参与了形成氨基咪唑环基团[16]。Skog等[17]在模拟体系中使用了14C标记的葡萄糖,并在产物MeIQx、4,8-DiMeIQx和IQx分子中检测到标记的碳原子,因而证明了葡萄糖是这些杂环胺的前体物。Chen等[18]发现单独加热肌酐没有检测到杂环胺的生成,而在150 ℃及以上温度加热葡萄糖和肌酐则可以生成AαC、MeAαC。Gibis等[19]以10种动物肉为原料做成馅饼,研究了前体物与杂环胺生成的关系,发现赖氨酸与PhIP呈高度显著相关(r=0.77,P<0.001),酪氨酸、苯丙氨酸、脯氨酸等也与PhIP呈显著正相关(r=0.47~0.56,P<0.05),相比之下,4,8-DiMeIQx、Norharman和Harman与任何游离氨基酸水平均无显著相关性;Norharman和Harman显示为葡萄糖含量的函数;PhIP与总肌酸(酐)和葡萄糖的物质的量比极显著相关(r=0.87,P<0.001),使用非线性回归(r=0.497,P<0.001)比线性回归(r=0.35,P<0.01)更好地描述了MeIQx中与总肌酸(酐)和葡萄糖的摩尔比关系。不同食物基质含不同比例、含量的前体物[20-22],在杂环胺生成的种类、含量上会有较大的差异[23]。
2.2 烹饪条件及食品基质其余组分对杂环胺形成的影响
除了前体物的影响,食品中杂环胺的生成还受到烹饪温度、时间和食品中其余组分如水分、脂肪、铁含量等因素的影响。Ishak等[24]通过在多个温度下利用模拟体系研究PhIP形成动力学发现:PhIP形成的温度和时间变化之间存在显著相互作用(P<0.05);苯丙氨酸模型系统在150 ℃开始形成PhIP,随着时间延长和温度的升高,PhIP含量显著增加(P<0.05),直到240 ℃加热8 min达到最高生成记录,为21.41 μg/mL,而后随加热时间的延长含量逐渐下降。Chiu等[25]关于杂环胺热稳定性的研究认为是高温导致了PhIP的分解。鄢嫣[10]通过主成分分析(PCA)发现,动力学曲线仅能分开直观反映各种杂环胺的形成规律,而建立的PCA模型很好地验证了高温短时焙烤样品与低温长时焙烤样品生成情况更加靠近;其中PhIP、4-OH′-PhIP、IQ[4,5-b]在250 ℃加热30 min和225 ℃加热35 min后含量开始下降。Lu等[26]通过使用植物油代替动物脂肪制作猪肉馅饼,发现植物油的类型会影响除IQ以外的杂环胺,并显著影响总杂环胺含量(P<0.05),同时对蛋白质氧化的影响显著(P=0.001);总杂环胺含量和TBARS(r=0.826,P<0.01)、羰基蛋白(r=0.778,P<0.01)之间呈显著正相关,进一步证实脂质氧化和蛋白氧化均参与了杂环胺的形成。郭海涛等[27]发现脂肪含量的增加会显著提高极性杂环胺的生成量(P<0.05),而对非极性杂环胺含量则无显著性影响。Lu等[23]的试验证明,与鸡肉相比,牛肉含有较高水平的非血红素铁,可以通过与羟基、过氧自由基发生反应来加速脂质氧化和美拉德反应进程,使得在对照样品中,牛肉丸的总杂环胺含量显著高于鸡肉丸(P<0.05)。因此,在不同报道中同种类食品杂环胺生成的差别,一定程度上受食品中其他组分的影响。
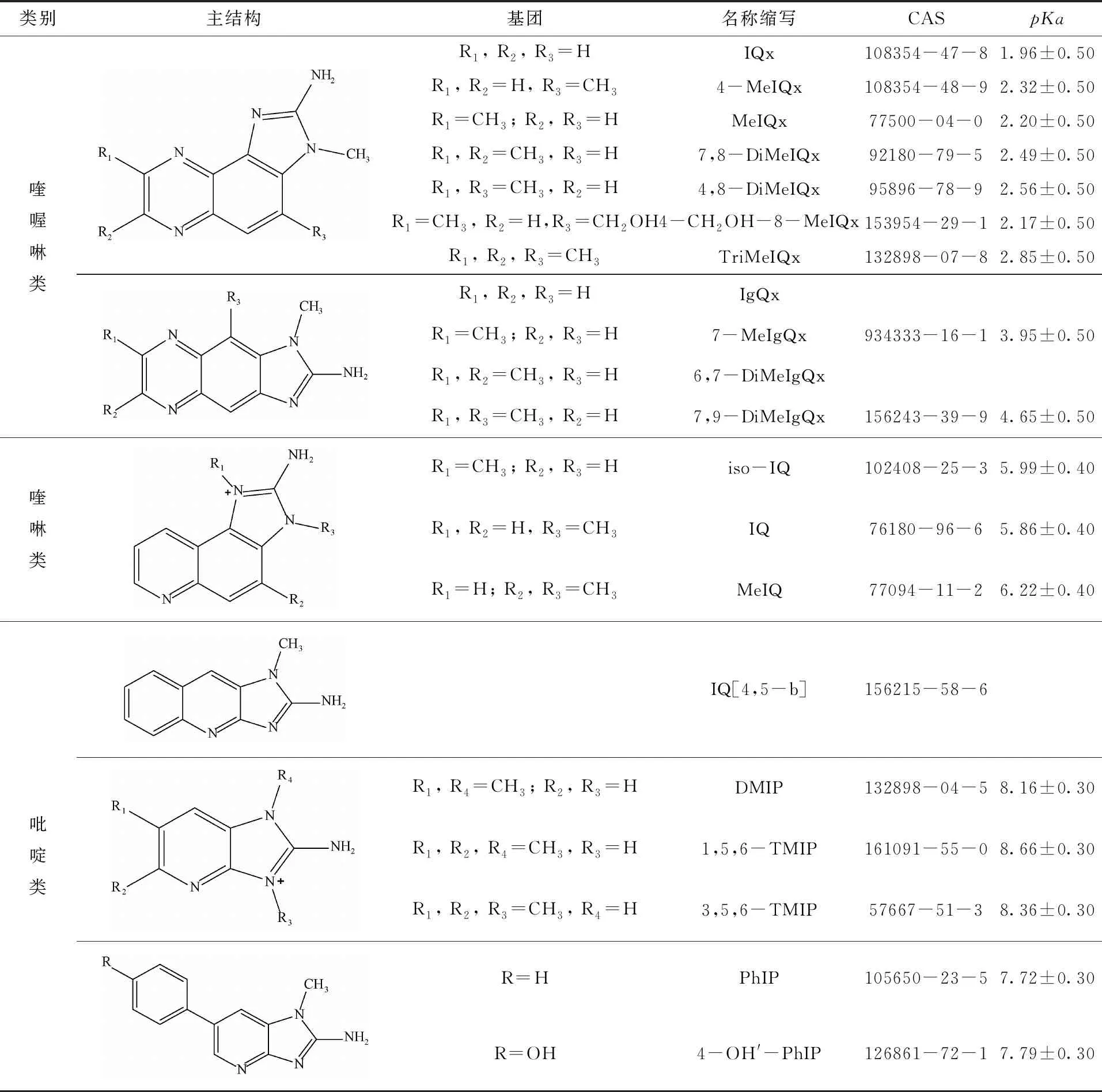
表1 氨基-咪唑-氮杂芳烃类杂环胺分类及理化性质
食品中杂环胺以游离态和结合态两种形式存在。Kataoka等[28]发现可以通过酸水解或酶解将PhIP从食品大分子化合物中释放出来,通过加热PhIP与白蛋白制备杂环胺与蛋白质的结合物,认为结合态杂环胺是在游离态杂环胺产生的基础上再与蛋白结合,后续在干热模型体系中发现PhIP-甘氨酸、IQ-甘氨酸、MeIQx-甘氨酸和Trp-P-1-甘氨酸加合物可在200 ℃加热10 min产生,过程如图1所示,其中PhIP-甘氨酸加合物最佳形成条件为200 ℃加热5 min,且加合物可在200 ℃下稳定保持至少20 min[29]。Chen等[30]发现在较低温度(小于200 ℃)时结合态杂环胺的种类比游离态杂环胺多;存在4种单一形式存在的杂环胺,6种以两种形式存在的杂环胺;因Harman、Norharman的主要贡献使得结合态杂环胺含量远远高于游离态杂环胺,而Harman、Norharman则是广泛存在于各类食品中的两类杂环胺[31],意味着在以往未区分杂环胺存在状态的研究中,高蛋白食品中杂环胺的实际含量比检测所得含量要高。

表2 氨基咔啉类杂环胺的分类及其理化性质
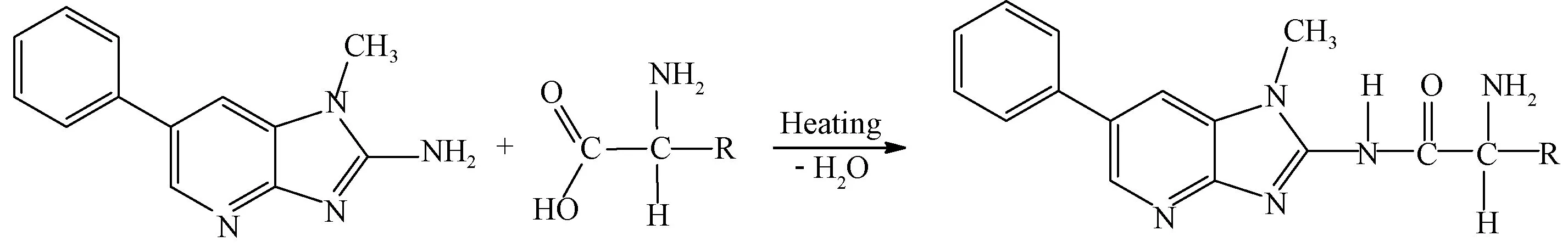
图1 PhIP与氨基酸加合物的形成[29]
3 杂环胺的抑制
3.1 新型加热方式对杂环胺产生的影响
过高的热加工温度和热处理时间的延长会不可避免地造成杂环胺含量的升高,但随着新型加热方式如微波加热、红外加热、过热蒸汽加热等方式的逐渐普及,研究显示新型加热方式使得杂环胺含量低于传统的加热方式。Suleman等[32]对比了炭烤、红外烧烤和过热蒸汽烧烤对羊肉饼加工过程中杂环胺的生成情况发现:炭烤组中存在9种极性杂环胺,总量为555.58 ng/g,红外烧烤存在7种极性杂环胺,总量为181.48 ng/g,过热蒸汽烧烤存在7种极性杂环胺,总量为30.67 ng/g;炭烤组中非极性杂环胺总量为426.06 ng/g,红外烧烤中非极性杂环胺总量为148.59 ng/g,过热蒸汽烧烤中非极性杂环胺总量为89.65 ng/g。值得注意的是,红外烧烤相对于炭烤和过热蒸汽烧烤极性杂环胺中引入了MeIQ,含量达到11.66 ng/g;而在非极性杂环胺方面,炭烤独有AαC,含量达到16.88 ng/g,过热蒸汽烧烤独有MeAαC,含量达到7.32 ng/g。Yao等[33]使用了3种木炭进行炭烤鸡腿,并与电烧烤、煤气烧烤进行了对比,其中,煤气烧烤杂环胺总量最高达30.38 ng/g,电烧烤杂环胺总量最低为12.43 ng/g。李利洁[34]利用模拟体系研究PhIP的生成规律发现:相对于传导密封加热最高生成量约2.45 ng/mL,使用微波回流加热和传导回流加热所生成的PhIP含量明显偏低,其中微波回流加热含量不到0.1 ng/mL。
3.2 添加外源物及食品加热前预处理对杂环胺的影响
自由基反应对杂环胺的形成起着重要作用[35],因此,预期抗氧化剂的加入能减少高蛋白食品中杂环胺的形成,但以往的报道存在与预期相反的情况。许多研究人员认为具有抗氧化特性的物质的抗氧化或促氧化作用可能因其结构、浓度、使用方法以及其余底物影响,而对杂环胺的抑制具有不同的表现。Mehr等[36]研究了肉豆蔻精油、生姜精油及其纳米乳液对烤牛肉饼中杂环胺的影响,试验中使用DPPH清除率评价了外源添加物的抗氧化能力,使用Minitab©19进行相关性分析(图2),r值变化范围0.936~0.986,显示DPPH清除率与杂环胺抑制率有较好的正相关性。但Nuray等[37]在研究中发现:与对照组相比,具有抗氧化能力的洋葱水提取物的使用导致了总杂环胺含量的增加。Salazar等[38]通过研究25种黄酮类化合物对PhIP的抑制作用时发现,芳香环位有两个羟基的酚类是最有效的抑制剂,烷基或羧基作为芳环中额外取代基存在会减弱多酚物对PhIP的抑制作用,引入额外羟基和氨基很大程度抵消了多酚的抑制作用,在赵磊等[39]的研究中同样印证了这一观点。鄢嫣[10]使用大豆分离蛋白、组织蛋白、淀粉等肉制品中常用配料研究对杂环胺形成的影响,发现这些配料的添加会促进杂环胺的生成,推测与蛋白高温加热后产生的游离氨基酸和羰基化合物有关,而当添加量达到10%时杂环胺含量开始降低,这与添加物的物理阻隔效应,限制杂环胺前体物的转移、接触有关。

图2 DPPH清除率与杂环胺抑制率相关性分析
Mehr等[36]将牛肉饼冻存90 d,然后加热检测杂环胺,发现随冻藏时间的延长,杂环胺的生成量逐渐下降,认为是冻藏减少了水分含量(P<0.05),从而限制了杂环胺前体物向食品表面的迁移,使得杂环胺生成量减少。张昆[22]在研究冻藏对肉制品杂环胺生成情况的影响时,重点研究了多次冻融对杂环胺生成的影响,试验发现猪肉、牛肉和鸡肉在多次冻融后结合态杂环胺含量都明显增加(P<0.05),推测与反复冻融导致的脂质、蛋白的氧化有关。
4 以热处理蛋白为模拟体系研究杂环胺的生成及抑制机理
热加工食品中蛋白含量、氧化程度、降解程度等往往和杂环胺的形成呈现相关性,但以蛋白为目标物系统研究杂环胺生成机理方面的研究比较少。随着科技发展和食品工业的变革,寻找肉类替代品解决肉制品供给不足、肉类相关的道德和环境问题成为一种可能,在零售和食品服务领域的全球食品市场中,肉类模拟食品行业正在迅速发展。目前的人造肉包含植物蛋白肉和细胞培养肉两个方面,其中植物蛋白产品已在蛋白质食品领域中占据一席之地,更容易被消费者接受[40]。植物蛋白肉的主要来源是大豆蛋白和小麦面筋蛋白,基于大豆蛋白的食品在加热情况下会有杂环胺的生成[31,41-42],因此未来对植物肉安全性方面的研究亟待加强[43]。本实验室在早先研究中发现干热大豆分离蛋白中含有多种杂环胺(图3),因此以蛋白为氨基酸的提供源,代替以往氨基酸-还原糖-肌酸(酐)模拟体系,研究蛋白制品杂环胺的生成机理更为合理。
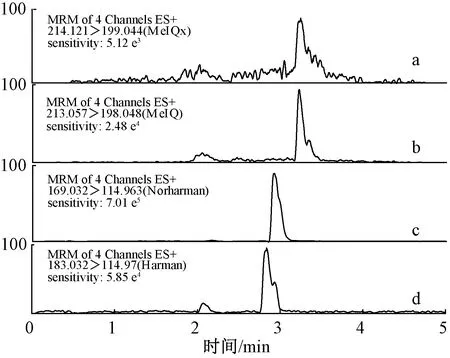
注:4个图的纵轴均表示强度/%。
杂环胺的生成直接受氨基酸含量、羰基化合物、自由基反应的影响,而蛋白在热处理过程中发生的系列复杂反应会影响上述杂环胺生成条件。利用共价交联形成多酚-蛋白复合物来稳定蛋白结构,提高蛋白抗氧化性,改善热处理过程游离氨基酸释放等方面的研究,对蛋白食品中抗氧化剂的加入对杂环胺生成抑制机理的解释提供了很好的参考依据。氨基酸侧链氧化是蛋白羰基形成的主要途径,并且是唯一被证实肉蛋白中羰基的形成机制[44],羰基含量与总巯基、自由氨基极显著负相关(P<0.01)[45]。为了减少具有一定毒性的人工合成抗氧化剂的使用[46],天然抗氧剂在食品中的应用得到广泛研究。多酚是广泛存在于植物体内具有抗菌、抗氧化、提高机体免疫力等多种生物活性的一大类天然产物,在食品中作为食品添加剂的应用也十分广泛。在蛋白体系中,多酚可以通过共价作用和非共价作用与蛋白结合改变蛋白的结构和性质。桑葚多酚可以提高猪肉干加工贮藏过程中蛋白氧化稳定性[47],低浓度绿原酸即可抑制羰基的生成,抑制蛋白氧化[48],Utrera等[49]的研究则显示天然抗氧化剂对肌原纤维蛋白羰基的影响受到多酚浓度、氧化条件、蛋白质靶标的影响。在植物蛋白方面,焦铭[50]的研究发现儿茶素-蛋白相互作用提高氧化蛋白消化后的营养特性,释放了更多的游离氨基酸。姜丽君[51]发现单宁可以结合小麦面筋蛋白上的自由氨基,与张慧芸等[52]在多酚-肌原蛋白方面的结果互相印证,表明多酚-蛋白作用机理在动物源蛋白和植物源蛋白方面具有共性,对杂环胺的产生具有潜在影响,但尚未有相关研究报道。
5 展望
研究高蛋白制品中杂环胺产生和抑制机理多使用真实食品热处理和氨基酸-还原糖-肌酸(酐)模拟体系,对杂环胺产生抑制方面的研究,除考虑引入新型健康加热方式外,多集中于研究添加天然抗氧化剂如富含多酚、黄酮类物质后对食物中杂环胺生成量的影响,但目前在杂环胺生成、抑制机理上研究解释仍有不足。真实食品热处理的结果在不同文献中有所出入,而使用现有的模拟体系推导所得结论不能很好地应用于真实食品中,因此,开发既接近真实食品基质,又能避免无关反应影响的模拟体系更有助于杂环胺抑制研究的实际应用。随食品工业的快速发展,植物肉或在不久的未来进入快速发展,意味着更多类型高蛋白食品被研发制造出来,持续加大对蛋白食品中杂环胺的产生及抑制研究将有利于食品工业的健康发展。
