2018年上海市活禽市场6株H9N2亚型禽流感病毒全基因进化分析
2021-05-20葛菲菲杨德全鞠厚斌周锦萍
葛菲菲,吴 杰,李 鑫,刘 健,杨德全,鞠厚斌,周锦萍
(1.上海市动物疫病预防控制中心,上海201103;2.扬州大学生物科学与技术学院,扬州225009)
禽流感病毒(Avian influenza virus,AIV)为单股负链RNA病毒,基因组由8个基因节段组成。H9N2亚型AIV是中国大陆家禽中最普遍的AIV之一[1],在家禽中稳定并持续存在,对养禽业造成严重损失和对人类健康构成严重威胁。其中G57基因型H9N2亚型AIV在2007年出现,并在2010年至2014年占据主导位置[2]。H9N2亚型禽流感病毒的6个内部基因节段是新型H7N9和H10N8亚型禽流感病毒的重配来源[3-4]。人H5N1、H7N9和H10N8亚型禽流感病毒的感染来源主要为活禽市场[5-7]。本研究对上海市活禽市场2018年6月分离到的6株H9N2亚型AIV进行全基因进化分析,为了解上海地区H9N2 AIV流行情况及其防控和候选疫苗选择提供有效的依据和参考。
1 材料与方法
1.1 病毒株 6株H9N2亚型AIV毒株由本实验室参照文献[8]分离保存,其详细背景见表1。
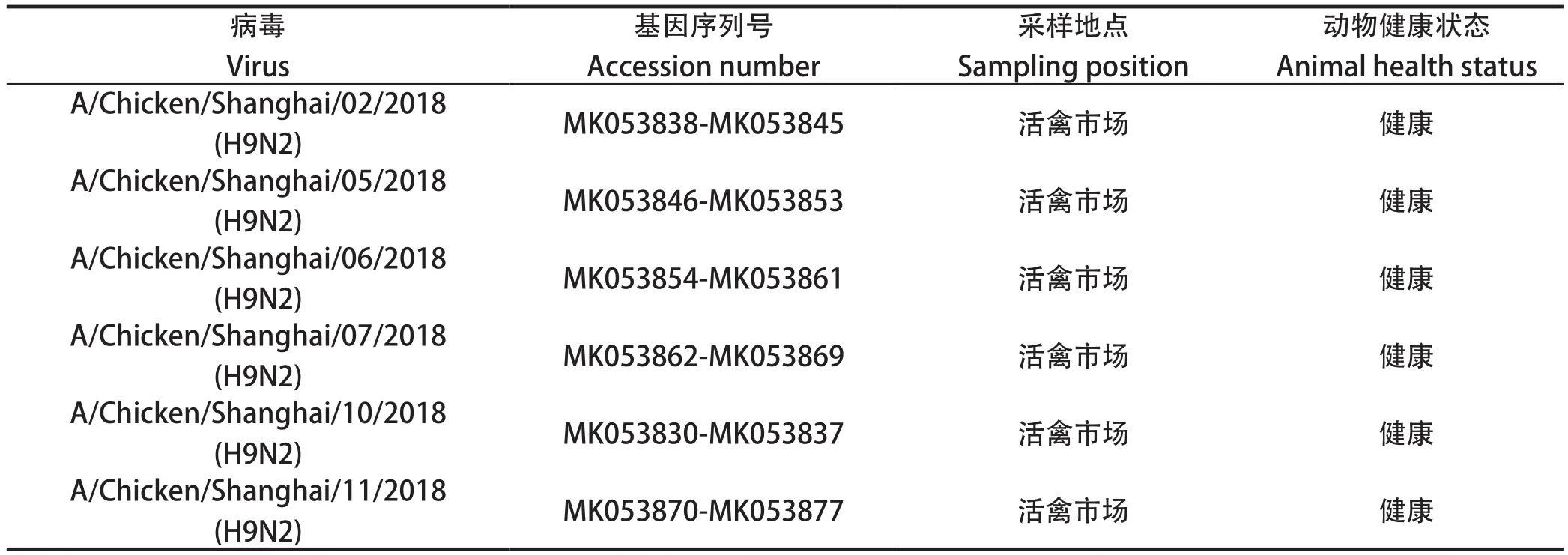
表1 本研究中6株 H9N2禽流感病毒毒株的信息Table 1 Information of the six H9N2 subtype Avian influenza virus isolates
1.2 实验动物及主要试剂 SPF鸡胚购自北京梅里亚维通实验动物技术有限公司;H9荧光RT-PCR试剂盒由深圳匹基生物工程股份有限公司提供;新城疫阳性血清、禽流感血凝素分型血清(H5、H9)由哈尔滨兽医研究所提供;AMV反转录酶等试剂均购自宝生物(大连)工程公司;特异引物由宝生物(大连)工程公司合成。
1.3 8个基因的扩增及序列分析 按照文献报道[9]的方法进行病毒RNA的提取和8个基因的全长扩增,测序由上海桑尼生物科技有限公司完成。参照GenBank中相关参考病毒株,利用Mega6.0软件进行序列的系统进化分析。
2 结果
2.1 全基因组同源性分析结果 6株病毒全基因片段的NCBI登录号为MK053830-053877。分离株HA基因ORF全长为1683 bp,编码560个氨基酸,核苷酸同源性为94.2%~99.3%,氨基酸序列同源性为93.9%~99.1%。分离株的HA基因与A/Duck/Y280/1997的HA基因核苷酸同源性为88.1%~99.3%,氨基酸同源性为89.7%~99.1%。分离株NA基因ORF全长为1401 bp,编码466个氨基酸,核苷酸同源性为96.0%~99.5%,氨基酸序列同源性为96.6%~99.6%。6株病毒内部基因M、NP、NS、PA、PB1和PB2核苷酸同源性分别为96.7%~99.7%、94.9%~99.7%、95.7%~99.4%、96.2%~98.9%、96.5%~99.9%、96.1%~99.9%。
2.2 HA基因关键位点分析 所有分离株的HA裂解位点均为RSSR↓GLF,为弱毒序列;受体结合位点的左缘(146 aa-150 aa)除了A/Chicken/Shanghai/07/2018为GTSKA,其余5株均为GTSTA;6株分离株HA唾液酸受体结合位点处氨基酸在191位为N(天冬酰胺),与经典的AIV国内分离株一致[10]。A/Chicken/Shanghai/07/2018在198位变异为A(丙氨酸),其余毒株均为T(苏氨酸)。A/Chicken/Shanghai/06/2018和A/Chicken/Shanghai/10/2018在202位和203位分别为R(精氨酸)以及F(苯丙氨酸),其余毒株在202位和203位分别均为L(亮氨酸)和Y(酪氨酸)。本研究中的毒株在受体结合位点的右缘(232 aa~237 aa)均为NGLMGR;此外本研究表明这6株毒株在HA基因226位点均为Leu,表明有向人类感染的嗜性[11-12](表2)。这6株病毒株的糖基化位点详见表3。
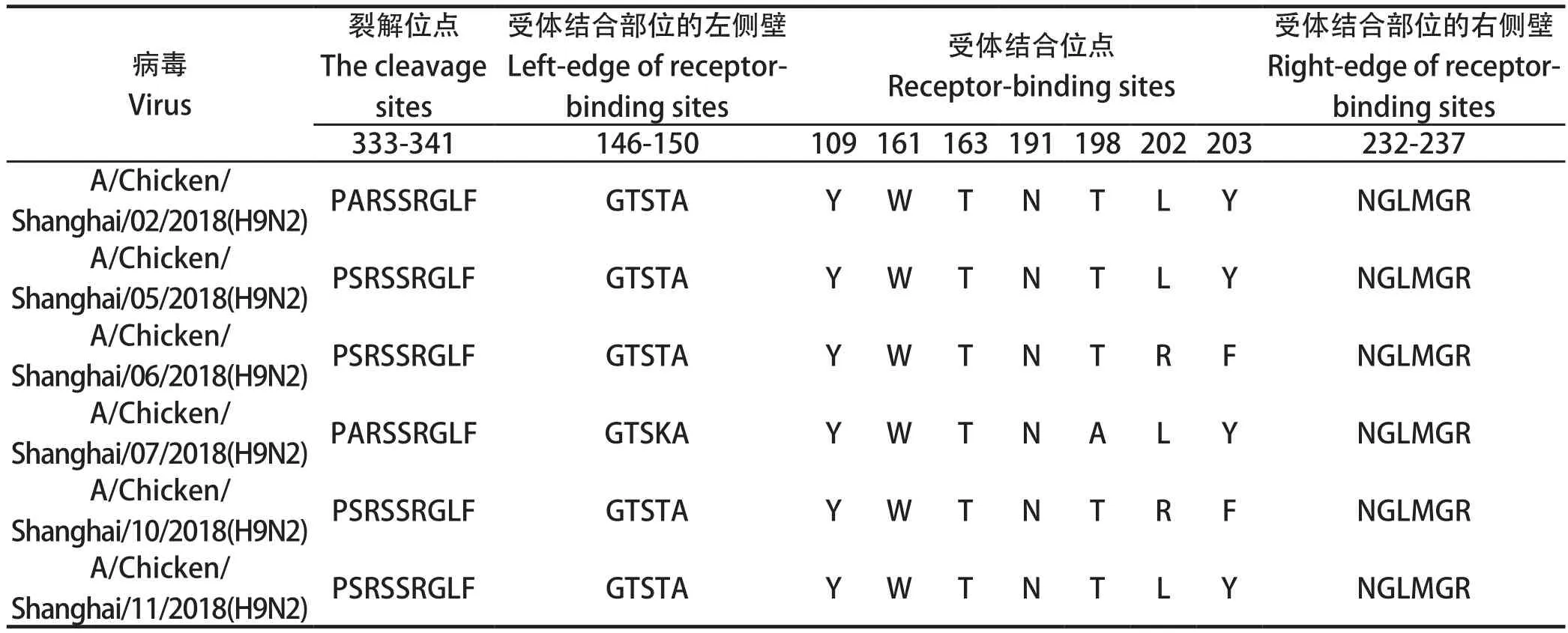
表2 H9N2分离株HA基因关键位点的比较Table 2 Comparison of several key sites of H9N2 subtype Avian influenza virus isolates

表3 分离株HA基因糖基化位点分析Table 3 Potential glycosylation sites of HA amino acid sequences in six H9N2 isolates
2.3 NA基因关键位点分析 对这6株分离株的红细胞结合位点,活性中心以及抗原决定簇进行了分析(表4),A/Chicken/Shanghai/10/2018(H9N2)和A/Chicken/Shanghai/11/2018(H9N2)在红细胞结合位点403位发生突变,在活性中心143位(同时为抗原决定簇的143位)发生突变,其余位点这6株病毒均保守。

表4 分离株NA氨基酸分析Table 4 Molecular characteristics of NA amino acid sequences in H9N2 isolates
2.4 HA基因遗传进化分析 利用Mega6.0软件对分离病毒株和参考病毒株HA基因的遗传进化关系绘制遗传进化树(图1)。结果显示,所有分离株均为Y280系。
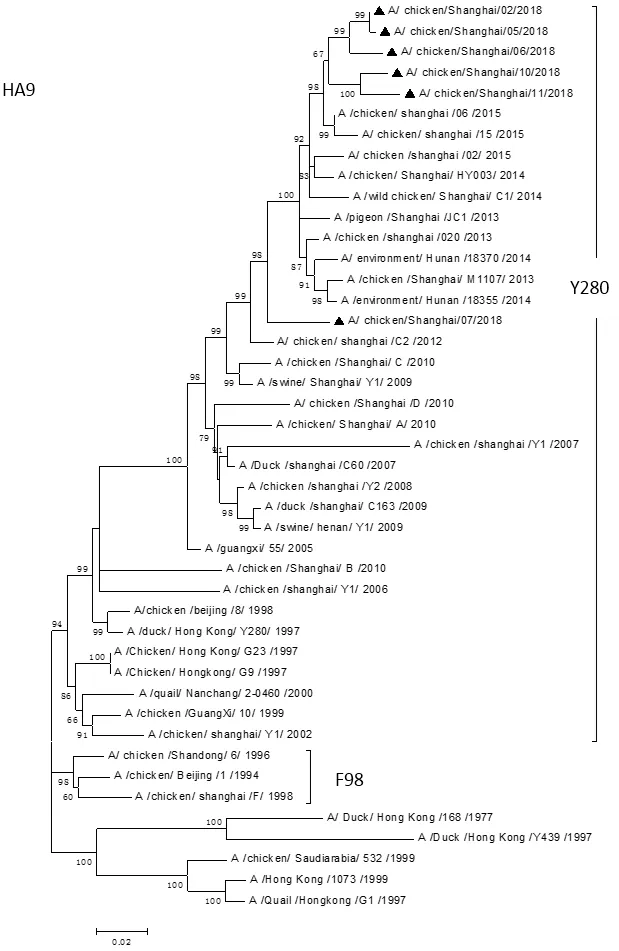
图1 6株H9N2亚型禽流感病毒HA的基因进化树Fig.1 Phylogenetic tree of HA genes of the six H9N2 subtype Avian influenza viruses
2.5 6株分离株的8个基因片段的重组模式 根据绘制的8个基因片段的遗传进化树分析(表5),6株分离株的重组模式完全相同,均为四重重组体,由Y280亚系、Unknown、G1以及F98重组而成,根据参考文献[13]判断,这6株病毒均属于G57基因型。
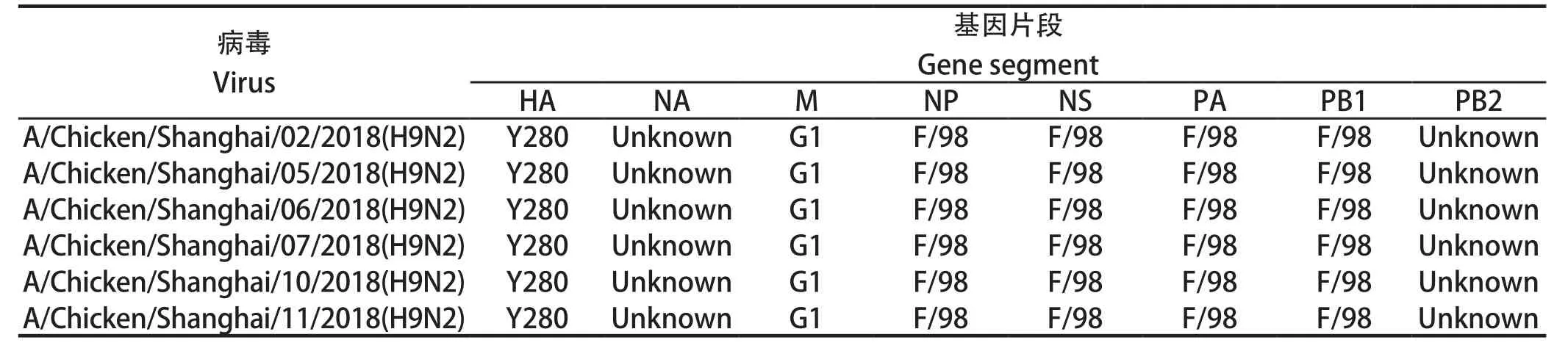
表5 8个基因片段的基因型组成Table 5 The genotypes of H9N2 subtype Avian influenza virus
3 讨论
对于AIV的监测始于20世纪70年代,然而流行病学的检测仅局限于养殖场、屠宰场及野生禽类中进行,直至1997年香港H5N1亚型AIV直接感染人并致人死亡事件的发生,活禽市场在AIV流行传播中的重要性被重视起来,对AIV流行情况的检测才逐渐开始[14]。本研究中发现6株分离株的HA基因第226位受体结合位点的氨基酸均为亮氨酸(Leu),已知HA基因中Q226L的突变能够与人类受体结合,呈现典型的人流感受体结合的特征[11,15];另外,本研究的6株H9N2亚型AIV基因裂解位点的序列均为P-S-R-S-S-R或P-A-R-S-S-R,并未出现多个碱性氨基酸的突变,有研究表明在HA裂解位点P5位置(316位点,H9编码)由丙氨酸到丝氨酸突变使得裂解位点氨基酸从模式-PARSSR-向-PSRSSR-的变异会增加病毒的毒力[16]。6株分离株HA蛋白第198、202和203位受体结合位点有所不同。A/Chicken/Shanghai/02/2018、A/Chicken/Shanghai/06/2018和A/Chicken/Shanghai/07/2018均有8个潜在糖基化位点,其余3个毒株存在9个潜在糖基化位点,有研究表明HA附加的糖基化位点和H9N2型AIV在家禽中的适应性之间存在联系[17]。
根据Zhu等[18]的研究,我们发现与2011-2014年中国分离的H9N2流感病毒相比,HA关键位点突变基本相似,对HA潜在糖基化位点的分析表明分离毒株与之前流行毒株相比未发生明显变化。本研究中的6株分离株NA上红细胞结合位点保持一致,与2011-2014年的毒株相比有位点发生突变;在NA活性中心只有A/Chicken/Shanghai/10/2018和A/Chicken/Shanghai/11/2018与2014年的2株分离株JS/TM71/14和AH/WB/14完全相同;与上述文献中的报道相比,NA上抗原决定簇主要的变异发生在367-370位和400-403位。
目前国内流行的H9N2亚型AIV的HA、NA虽然以CK/Bei-like分支基因型为主,但其内部基因经常被其他分支的基因片段所取代,出现越来越多的重组基因型。Gao等[3]研究表明,G57基因型在2007年出现,其流行率在2009年剧增,并自2010年后在中国占主导地位。本研究分离的6株H9N2亚型AIV均属于G57基因型,表明上海地区在2018年还是以G57基因型为主。综上所述,目前上海市活禽市场H9N2亚型AIV虽然以G57基因型为主,但其受体位点和糖基化位点等呈现多样化,一些关键位点的突变有可能引起感染性和传播力的增强,以上发现都值得进一步研究。同时H9N2亚型AIV内部基因为其他亚型AIV提供内部片段,加大了对人类健康的威胁,所以应该对其监测防控工作给予重视,有效防止新的疫情的发生。
