寨卡病毒非结构蛋白1对小鼠免疫应答的影响
2021-05-14荣小荣杨延鹏王垣芳
荣小荣 杨延鹏 赵 丹 王垣芳
1 滨州医学院药学院 山东 烟台 264003;2 聊城市第四人民医院 山东 聊城 252000
近几年来,由寨卡病毒(Zika virus,ZIKV)感染引起的ZIKV病在全世界诸多地区暴发流行[1-2],引起全球广泛关注。ZIKV感染除了引起发热、皮疹、关节疼痛、无力等症状外,还会对感染者的神经系统造成损害,孕妇感染可能导致胎儿畸形[1,3-5]。目前对于ZIKV病尚无特效的抗病毒药物和成熟的疫苗面市[6-8],因此研发特效的抗ZIKV药物和有效的ZIKV疫苗是亟待解决的问题。
ZIKV非结构蛋白1(Non-structural protein 1,NS1)在病毒复制、致病机制及免疫逃逸中起着关键作用,可诱导机体产生抗体,是该病毒感染的主要抗原[9]。有研究发现,采用ZIKV-NS1 DNA疫苗候选物接种于小鼠可使小鼠免受ZIKV感染,提示NS1单独使用可能具有保护机体免受ZIKV感染的作用[10]。本实验采用pET28a-ZIKV-NS1原核质粒表达得到重组NS1,并将其接种于小鼠,通过检测小鼠血清IgG抗体、淋巴细胞亚群及细胞因子观察其对小鼠免疫应答的影响,探讨NS1作为ZIKV疫苗的潜力,为ZIKV疫苗的研发提供新的思路。
1 材料与方法
1.1 材料
1.1.1 菌种及质粒 大肠埃希菌E.coli Rosetta(DE3)为实验室留存;感受态细胞DH5α购自天根生化科技(北京)有限公司;原核质粒pET-28a购自美国Novagen公司。
1.1.2 主要试剂 限制性内切酶BamHI/HindIII购自英国NEB公司;流式细胞术检测相关抗体(CD3+-APC/Cy7、CD4+-FITC、CD8+-PerCP、IFN-γ -APC、IL-4 -PE及IFN-γ测试盒(2210006)及IL-4(2210403)测试盒(达科为生物技术有限公司);Anti-ZIKV Virus NS1 antibody[B4](Abcam,批号为ab218546);HRP标记兔抗鼠IgG(二抗)(北京润泽康生物科技有限公司,批号为C2210);BCA蛋白浓度测定试剂盒(碧云天生物技术有限公司,批号为P0012)。
1.1.3 主要仪器 PCR仪(MyCyclerTM),Bio-Rad公司;核酸电泳仪(DYY-III-6B),北京六一仪器;凝胶成像系统(AlphalmagerTM 2200),Alpha公司;流式细胞仪(LSRFortessa),BD公司。
1.1.4 实验动物 6~8周龄SPF级雌性Balb/c小鼠,体质量为16~20 g,购自青岛派特福德白鼠养殖专业合作社,合格证号为SCXKL(鲁)20140007。
1.2 方法
1.2.1 ZIKV-NS1特异性引物的设计 通过基因合成得到ZIKV-NS1基因(Gene-Bank序列号KU963796.1),运用Primer 5.0软件设计上下游引物,在上下游分别插入酶切位点BamHI和HindIII,片段NS1全长1 056 bp,引物信息如下:
ZIKV-NS1-F:5′-CGCGGATCCATGGATGTGGGGTGCTCGGTGGA-3′,
ZIKV-NS1-R:5′-CCCAAGCTTTTATGCAGTCGACCATTGACCTT-3′。
1.2.2 PCR扩增目的片段 以ZIKV-NS1基因为模板,ZIKV-NS1-F/ZIKV-NS1-R为引物,进行PCR扩增。反应条件为:98℃预变性5 min;98℃变性30 s,68℃退火30 s,68℃延伸1 min,循环30次;68℃延伸10 min后结束;16℃存放。将得到的PCR产物上样于凝胶电泳仪进行跑胶得到目的条带,对目的条带进行切胶回收,再应用胶回收试剂盒回收目的片段。
1.2.3 pET28a-ZIKV-NS1构建与鉴定 应用限制性内切酶BamHI和HindIII对质粒和目的片段进行双酶切,酶切后的目的片段和质粒进行回收,用T4 DNA ligase连接,得到重组质粒(图1),提取重组质粒后送测序并鉴定,确定重组质粒为pET28a-ZIKV-NS1,将质粒转化至表达菌感受态细胞BL21(DE3)。

PT7.T7启动子;His6.多组氨酸标签;TT7.T7终止子。图1 ZIKV-NS1基因质粒构建示意图
1.2.4 ZIKV-NS1表达、纯化及鉴定 将鉴定正确的阳性菌落接种于LB培养基,震荡培养至菌液在600 nm处的吸光值在0.4~0.6之间,加入IPTG(终浓度为1 mmol/L)诱导NS1的表达,在20℃下继续培养18 h。离心收集菌体后用磷酸缓冲盐溶液(PBS)重悬,之后应用超声破碎菌体收集沉淀得到目的蛋白。对于包涵体蛋白采用8M尿素变性,透析复性的方法得到上清,因目的蛋白偶联His6标签,应用亲和层析法分离蛋白,将上清与柱子充分结合,经镍柱多次亲和挂柱及梯度咪唑洗脱后得到目的蛋白,将得到目的蛋白经BCA蛋白浓度定量试剂盒测定终得率约5 mg/L菌液,取样用于SDS-PAGE分析及Western Blot鉴定。最后,将目的蛋白溶解于磷酸缓冲盐溶液(PBS)中,分装后冻存于-80℃冰箱保存备用,避免反复冻融。
1.2.5 ZIKV-NS1小鼠免疫及标本采集 将10只小鼠随机分为空白对照组和实验组,每组5只。给空白对照组小鼠肌内注射PBS(100 μL/只),实验组小鼠肌内注射NS1(20 μg/只)进行免疫,每隔14 d注射1次,共注射3次。第1次和第2次注射1周后行内眦部取血,第3次注射1周后行摘眼球取血,将所采血液37℃静置2 h后以5 000 r/min离心5 min制得血清,每只小鼠均取20 μL血清进行效价检测。上述实验重复3次,总共使用小鼠30只。
1.2.6 小鼠血清IgG效价检测 采用酶联免疫吸附法(ELISA)测定,以PBS组为空白对照组,用纯化的ZIKV-NS1包被酶标板(终浓度为1 μg/mL),每孔中加入100 μL稀释好的蛋白溶液,4℃包被过夜,PBST洗板2~3次,每次300 μL,5%脱脂牛奶37℃封闭2 h备用。提前将采集好的小鼠血清用含有1%脱脂牛奶的稀释液1∶32倍起进行2倍梯度稀释,用稀释后的血清作为一抗进行孵育,兔抗鼠IgG抗体(HRP标记)为二抗,3次洗板后进行TMB显色,应用2M硫酸50 μL /孔作为终止液终止显色。应用酶标仪在450 nm处检测吸光值,相同稀释倍数下以实验组数据大于空白对照组数据2.1倍以上为阳性结果,以阳性血清的最高稀释倍数表示抗体效价。
1.2.7 脾脏CD4+、CD8+细胞数量及IFN-γ、IL-4分泌水平检测 将小鼠摘眼球取血后,取出脾脏加入淋巴细胞分离液,置于200目尼龙网上研磨,离心后分离得到淋巴细胞。将空白对照组(PBS)与实验组(NS1)均分为两组,第1组细胞不加入NS1直接应用流式细胞仪检测,检测出1万个细胞中CD4+、CD8+细胞数量及分泌IFN-γ、IL-4的细胞数量,分别定义为空白对照组(PBS)-未刺激(组1)和实验组(NS1)-未刺激(组2)。同时利用24孔板进行细胞培养,将第2组细胞每孔约5×105个细胞进行铺板,每孔加入20 μg NS1,每孔额外加入monensin 2 μmol/L(蛋白转运抑制剂),37℃孵育4 h后应用流式细胞仪检测1万个细胞中相同淋巴细胞亚群和相同细胞因子的细胞数量,分别定义为空白对照组(PBS)-刺激(组3)和实验组(NS1)-刺激(组4)。应用Flowjo7.6软件分析流式数据,测定CD4+、CD8+细胞数量及分泌IFN-γ、IL-4的细胞数量占样本中所有细胞的百分比。之后应用酶联免疫斑点测定法(ELISPOT)测定NS1特异性分泌IFN-γ、IL-4的细胞数量。
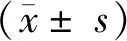
2 结果
2.1 ZIKV-NS1基因PCR扩增与重组质粒鉴定 以合成序列为模板PCR扩增后跑胶出现条带约为1 056bp,与合成目的序列大小相同。将质粒与目的片段进行双酶切,酶切后用T4 DNA ligase连接,得到重组质粒,对重组质粒进行双酶切验证,可见2条明显条带,大条带约为5 369 bp,与质粒大小相当;小条带约为1 056 bp,与目的序列大小相当,见图2。

A. ZIKV-NS1基因PCR扩增;B. pET28a-ZIKV-NS1质粒双酶切鉴定。M. DNA标志物;A1. ZIKV-NS1基因PCR产物;B1. 重组质粒双酶切鉴定。图2 ZIKV-NS1基因PCR扩增与重组质粒鉴定
2.2 重组NS1的表达纯化与鉴定 纯化所得蛋白经SDS-PAGE分析,在44 kD处有单一条带,经Western Blot 鉴定能与Anti-ZIKV-NS1抗体特异性结合,所得蛋白为重组ZIKV-NS1,单一条带用Image J软件进行灰度比较分析,灰度值高于85%,表明纯化得蛋白单一。蛋白经Western blot 鉴定能与Anti-ZIKV-NS1及His标签抗体特异性结合,所得蛋白为重组ZIKV-NS1,见图3。

A. ZIKV-NS1的SDS-PAGE分析;B和C. 重组ZIKV-NS1的Western blot 鉴定。M. 蛋白分子质量标准;1. 纯化的目的蛋白。图3 重组NS1的表达纯化与鉴定
2.3 小鼠血清IgG效价 空白对照组3次注射PBS后分别检测血清IgG效价,结果显示抗体效价水平均较低,3次结果均无明显变化。实验组小鼠初次NS1免疫后血清IgG抗体效价较低,第2次和第3次免疫后IgG效价明显升高,第3次免疫后IgG效价可达1∶18 000。实验组小鼠第2次和第3次免疫后IgG效价显著高于空白对照组,P<0.001,见图4。
2.4 小鼠脾脏CD4+、CD8+细胞数量及IFN-γ、IL-4分泌水平 实验组(NS1)-刺激(组4)CD4+、CD8+细胞数量及IFN-γ、IL-4的分泌水平均上升,与实验组(NS1)-未刺激(组2)比较,P<0.01;实验组(NS1)-刺激(组4)CD4+、CD8+细胞数量及IFN-γ、IL-4的分泌水平上升,与空白对照组(PBS)-刺激(组3)比较,P<0.01,见表1,图5、6。

与空白对照组相比,***P<0.001。图4 小鼠血清IgG效价
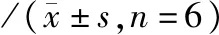
表1 小鼠脾脏CD4+、CD8+细胞数量及IFN-γ、IL-4分泌水平

图5 小鼠脾脏CD4+、CD8+细胞数量及IFN-γ、IL-4分泌水平

与空白对照组(PBS)-刺激比较,***P<0.001。图6 小鼠脾脏CD4+、CD8+细胞数量及IFN-γ、IL-4分泌水平
2.5 小鼠脾脏IFN-γ、IL-4分泌型细胞数量 实验组(NS1)小鼠脾脏IFN-γ分泌型T细胞斑点数量为每百万个细胞约135个,IL-4分泌型T细胞斑点数量为每百万个细胞约25个,空白对照组(PBS)无斑点,二者比较,P<0.001,见图7。

与空白对照组(PBS)比较,***P<0.001。图7 小鼠脾脏IFN-γ和IL-4分泌型细胞数量
3 讨论
ZIKV的非结构蛋白NS1是ZIKV感染的主要抗原,在病毒复制、致病机制和免疫逃逸中起着关键作用[9]。对于NS1的研究需选择适宜的蛋白表达系统,目前常用的有大肠杆菌表达系统和杆状病毒表达系统。杆状病毒表达系统周期长哺乳动物细胞表达系统表达量低、成本高[11-16],而大肠杆菌表达系统具有遗传背景清楚、周期短、表达量高、成本低等优点,因此本实验采用大肠杆菌原核系统诱导表达NS1,经SDS-PAGE分析及Western Blot鉴定表明重组NS1成功。
人体免疫应答包括体液免疫和细胞免疫。体液免疫是以B细胞产生抗体来达到保护目的,因此对抗体效价的测定是检查体液免疫的主要指标,其中IgG抗体可发挥持久的保护作用,是动物机体抗感染的主力。细胞免疫是T细胞借由释放淋巴因子而发挥免疫力的免疫,而T细胞功能取决于T淋巴细胞总值(CD3+细胞)及其亚群(CD4+细胞、CD8+细胞)的相对组成[17],正常情况下,亚群间互相拮抗达成平衡。因此T淋巴细胞计数及其亚群和细胞因子等是判断机体细胞免疫水平的重要指标。
IgG抗体是人体最重要的抗病原微生物抗体,能与病毒抗原结合,从而使病毒失去感染细胞的能力,且能够激活补体效应,诱导被感染细胞死亡,使其中的病毒也随之被清除[18-19]。本实验结果显示,给小鼠肌内注射NS1免疫 2次和3次后IgG效价升高,且随免疫次数的增加而升高,表明NS1可刺激机体产生IgG抗体,具有激活体液免疫的作用。
CD4+细胞、CD8+细胞及IFN-γ、IL-4等细胞因子在机体细胞免疫方面起着关键作用,是目前筛选病毒免疫原蛋白的主要指标[17,20]。当机体受到病毒感染时,CD4+细胞可辅助CD8+细胞参与机体细胞免疫应答,清除体内被病毒感染的异常细胞从而保护机体正常组织细胞免遭病毒侵袭,两个亚群比例可因此发生改变。机体免疫系统可通过细胞因子进行调节,IFN-γ、IL-4等细胞因子是免疫调节的关键物质之一[20]。其中IFN-γ是Th1细胞的代表性细胞因子,是一种高效的抗病毒生物活性物质,具有广泛免疫调节作用,包括诱导多种抗原提呈细胞表达MHC-I/II分子,活化单核、巨噬细胞、中性粒细胞和NK细胞,促进Th1细胞发育和抑制Th2细胞活化与增殖,刺激B细胞产生的抗体类型向调理素方向转变[21]。IL-4是Th2细胞的代表性细胞因子,可刺激B细胞和T细胞增殖,在调节体液免疫和适应性免疫中起关键作用[22]。本实验结果显示,NS1能使小鼠脾脏CD4+和CD8+细胞数量增加,提示NS1具有激活细胞免疫的作用;NS1能使IFN-γ、IL-4分泌水平升高,且NS1特异性IFN-γ分泌型T细胞斑点数量较IL-4的细胞斑点数量升高更明显(分别为每百万个细胞约为135和25个),提示ZIKVNS1促进Th1/Th2细胞平衡向Th1细胞偏移,以Th1型细胞介导的细胞免疫为主。
近年来对于NS1免疫原的研究多侧重于体液免疫[15-16],而对于NS1对细胞免疫的影响研究甚少,本实验通过重组蛋白NS1从体液免疫和细胞免疫两个方面进行了研究。前期Grubor等[10]研究发现NS1-DNA疫苗候选物可通过增强小鼠细胞免疫功能从而抵抗ZAKV感染,与本研究中NS1可激活小鼠细胞免疫是一致的。但目前多数国家核酸疫苗研发技术基础薄弱,安全性未知,且在高等动物体内难以诱导较强的免疫应答[23],而重组蛋白疫苗研发技术已较成熟,且安全、高效、投入生产和应用的效率会更高。
ZIKV属于黄病毒属,黄病毒属病毒基因组编码至少三种结构蛋白(衣壳蛋白C、膜蛋白M和包膜蛋白E)和七种非结构蛋白,其中E蛋白是病毒的重要抗原成分[24]。目前对于黄病毒疫苗的研发主要是应用黄病毒E蛋白激活体液免疫[7-9,25]。某些黄病毒E蛋白相似的基因序列和结构同源性可导致高度的抗体交叉反应性,并导致感染的抗体依赖性增强(antibody-dependent enhancement,ADE)[26]。有研究显示,以传统黄病毒E蛋白构建的重组腺病毒载体疫苗中插入NS1基因能够明显增强疫苗的保护效力[27-29],且黄病毒NS1上存在多个特异性的抗原表位,由此推测,NS1可能会通过激活机体免疫系统而增强E蛋白载体疫苗的保护效果,且NS1不表达于病毒粒表面,不会引起ADE[28,30-31]。
本实验通过原核表达系统表达并纯化得到ZIKVNS1,并使用NS1对小鼠进行免疫,结果显示NS1能使小鼠血清IgG效价显著升高、脾脏CD4+和CD8+细胞数量及IFN-γ、IL-4分泌水平升高,提示NS1对小鼠体液免疫和细胞免疫均具有激活效应,NS1具有成为ZIKV疫苗的可能性;与近年来ZIKV NS1-DNA疫苗的研发思路不同,本研究可能为ZIKV疫苗和黄病毒疫苗的研发提供了新的思路。
