华北落叶松人工林地表处理措施对当年幼苗密度的影响*
2021-04-10王彦辉余治家王正安贾宝光
佘 萍 曹 兵 王彦辉 余治家 王正安 马 杰 贾宝光
1.宁夏农林科学院固原分院 固原 756000; 2.宁夏大学农学院 银川 750021; 3.中国林业科学研究院森林生态环境与保护研究所 国家林业和草原局森林生态环境重点实验室 北京 100091)
自然下种萌发是森林(尤其针叶林)天然更新的最主要途径,有利于降低造林成本并形成近自然林。如何对其深入理解和科学利用一直是森林经营、森林生态、森林恢复等学科的关注热点(Puettmannetal., 2005)。在种子落地、萌发、幼苗形成和幼树定居的各更新阶段,都会受到许多自然和人为因素干扰(陈永富, 2012; 程瑞梅等, 2018),包括树种特征、种子大小与品质(Bonfil, 1998),也包括立地环境(杨莎, 2014; 程瑞梅等, 2018)、林分结构(曹怡立等, 2019)、枯落物覆盖(李进等, 2020)、种子库动态(Carltonetal., 1998),还包括干扰类型、林窗特征、天然更新方式及过程等(汤景明等, 2005; 李小双等, 2007),且主要影响因素随环境和树种等变化而不同。研究表明,枯落物层过厚会阻碍幼苗根系进入矿质土壤,影响幼苗抗旱能力(Scariot, 2000; Essery, 1998); 土壤理化性质(pH和C、N、P含量等)对乔木更新也有重要影响(Muscoloetal., 2007); 林窗大小通过影响林内光环境影响幼苗生长和存活(Huthetal., 2006)。虽然单纯封禁也能促进天然更新,但其作用有限,有时必须人工更新或人工促进天然更新(董伯骞等, 2014),如间伐制造林隙(Myersetal., 2000)、落种前扰动土壤(Dobrowolska, 2010)或通过计划烧除(Lucas-Borjaetal., 2016)排除灌木竞争(Robertsetal., 2017)。目前,关于天然更新的研究主要集中在天然更新过程和影响因素方面,对一些重要树种的人工促进天然更新研究还相对不足。
华北落叶松(Larixprincipis-rupprechtii)是我国北方的主要造林树种,宁夏六盘山地区于20世纪60年代从山西引入,现人工林面积占当地人工林总面积的80%以上,发挥着重要生态功能。研究发现,宁夏六盘山华北落叶松幼苗密度在主要森林类型中最低(金红喜等, 2009),其可持续经营受制于天然更新的实用不足和缺乏人工促进天然更新的实用技术。目前,六盘山地区华北落叶松林研究主要集中在生态水文方面(王云霓等, 2015; 王艳兵等, 2017; 韩新生等, 2019),天然更新特征和影响因素研究还很少(杨占彪, 2009);有限的研究主要在河北省和山西省,如冀北山地退化华北落叶松人工林封禁后的林隙内更新密度高于林冠下,幼苗在开敞度为0.4~0.5的林隙内生长最好(董伯骞等, 2014)等; 更新幼苗数量随林隙面积增大而增加,并在160~180 m2时最大(盖力岩等, 2014); 在河北塞罕坝地区,土壤-枯落物因子是影响更新的关键(张树梓等, 2015),土壤pH和全磷含量也显著影响幼苗密度(孙国龙等, 2017)。在山西关帝山,华北落叶松纯林的更新苗量随林分密度增大而减少(杨莎, 2014); 更新苗呈聚集分布,聚集程度随林分密度增大而升高(王旭刚, 2018)。综合来看,虽有一些华北落叶松林天然更新和影响因素的调查研究,但还需在控制条件下研究、确定主要影响因素,特别是发展人工促进天然更新的实用技术,以指导生产实践。
考虑到华北落叶松天然更新可能在不同环节受多种因素影响,包括种子数量和质量、动物取食种子、枯落物层阻隔幼苗根系在土壤中发育、林冠遮光、林下植被竞争等,本研究在宁夏六盘山区设计不同生境森林样地的多种地表处理措施,人工撒播一定数量的种子,比较各种地表处理措施对当年幼苗密度的影响,以期找到华北落叶松天然更新的主要限制因素,发展人工促进天然更新的实用技术,为改善华北落叶松林天然更新和林分结构提供技术支撑。
1 研究区概况
研究区位于我国西北黄土高原中西部的宁夏六盘山(106°09′-106°30′E,35°15′-35°41′N),地处半湿润-半干旱区过渡带,年均气温6.1~6.8 ℃,年降水量450~670 mm,是黄土高原主要水源地。土壤以山地灰褐土为主,占总面积的94.44%。土层平均厚度30~60 cm,薄厚分布不均,土壤有机质含量较高。天然林主要树种有白桦(Betulaplatyphylla)、辽东栎(Quercusliaotungensis)、华山松(Pinusarmandii)、椴树(Tiliaamurensis)、榆(Ulmuspumila)和小叶杨 (Populussimonii)等,人工林主要树种有华北落叶松、油松(Pinustabulaeformis)和云杉(Piceaasperata)等。
2 研究方法
2.1 样地设置
选择在六盘山的东部(彭阳县古城镇挂马沟村)、西部(隆德县峰台乡清凉河水坝)、南部(泾源县新民乡大雪山)、北部(泾源县大湾乡杨岭村水沟)和中部(泾源县惠台乡米缸山二台)5个试验点,对每个试验点的华北落叶松人工近熟林(30~40年生),按林冠郁闲度不同,选择树冠下(正常生长树木的冠下)、林隙(林窗,指树木死亡或移除后形成的林冠层空隙,面积300~500 m2)、林中空地(林中小片无林地段,面积1 500~2 000 m2)和林缘4类样地,并在每个试验点林缘附近选择2000年后退耕还林栽植的华北落叶松幼龄林(15~20年生)样地,共5类生境,每类生境设置1块12 m × 20 m样地。整个试验共设置25块样地,不同试验点的各生境类型基本情况见表1。
在每块样地,设计了5个处理: 割草(用割草机全部割掉地上2 cm以上草本,并清除枯落物未分解层)、开沟(平行于等高线用锄头开深10 cm、宽15 cm的沟,沟间距60 cm,露出矿质土壤)、挖穴(深20 cm,上口直径20 cm,穴间距20 cm,露出矿质土壤)、清枯(全部清除草灌及整个枯落物层,露出矿质土壤并松土)和对照(无任何地表处理)。 每种处理样方的面积均为4 m×4 m,3次重复,采用完全随机区组设计,每个区组平行于等高线排列,每个区组间的走道宽0.5 m,区组周围的保护行宽0.5 m。虽然开沟和挖穴处理的目的都是为了露出矿质土壤以使种子直接在土壤上萌发,但整地的深度和面积比例及导致的施工难度和成本差别均较大,而且种子被滑落土壤埋藏深度不同也可能影响种子萌发和幼苗发育。
此外,在5个试验点的近熟林和退耕幼龄林的林冠下,4 m×4 m的处理样方中间选择1块1 m×1 m的小样方,播种后覆盖孔径0.5 cm的尼龙网,尼龙网距地面20 cm,以防动物取食种子,小样方外播种后不覆网,观察覆网对提高幼苗密度的作用。
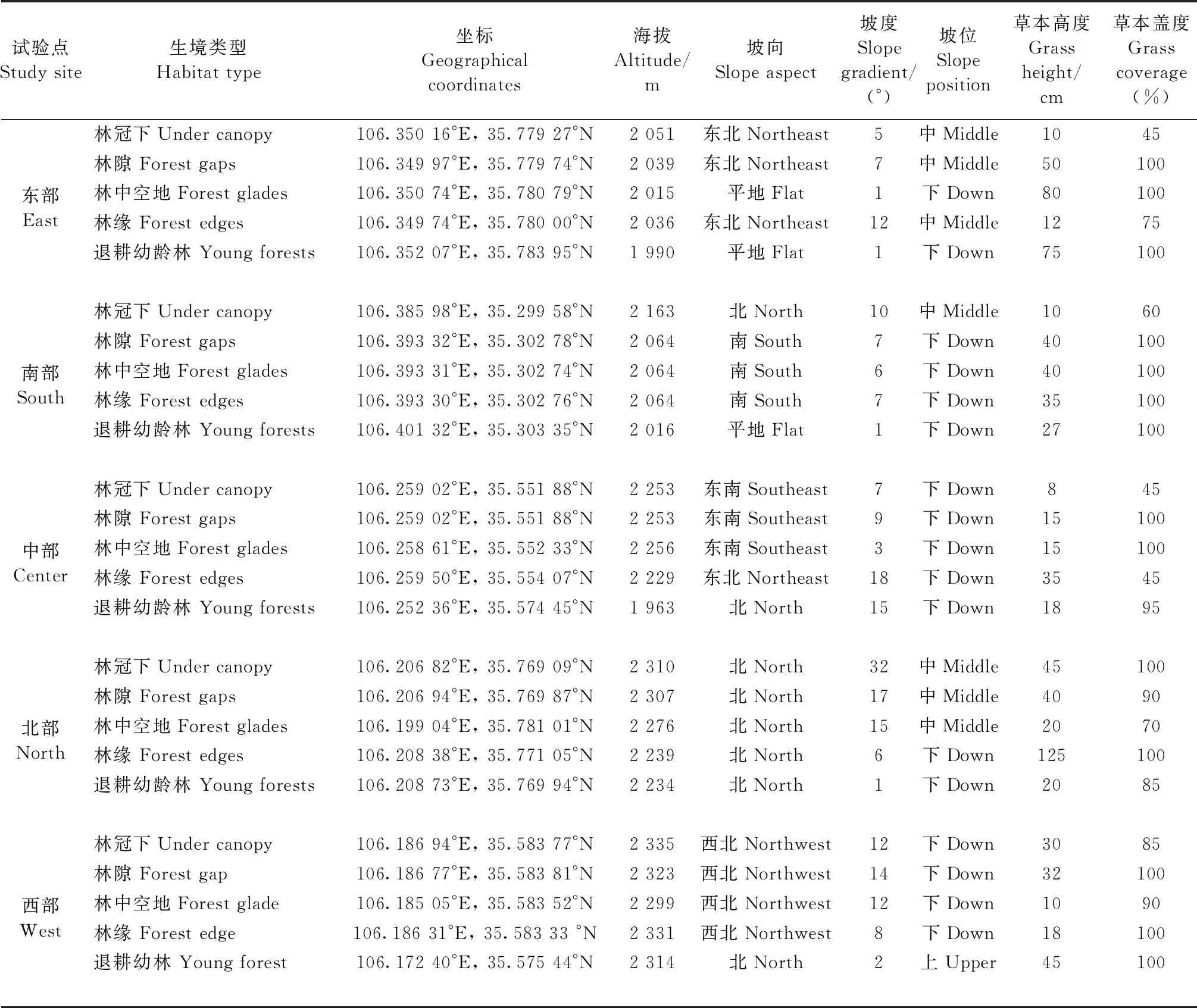
表1 样地概况
2.2 种子撒播
2018年10月25—30日完成所有样地的地表处理,并人工模拟自然下种,在地表撒播种子(100粒·m-2)后未覆土或枯落物。种子来自六盘山华北落叶松种源地(山西省忻州市繁峙县),发芽率为31.4%。试验前调查所有试验点和各类生境,均无天然更新幼苗,故认为天然下种对本试验无影响或影响极小,可忽略不计。
2.3 野外调查与数据分析
华北落叶松出苗期较长,在六盘山为5月初到6月中旬,播种后第2年(2019年6月20—30日)人工计数各样方的幼苗密度,对比不同处理的差异。
采用SPSS 19.0软件进行单因素方差分析和Duncan多重比较,运用Excel 2010和Graphpad prism 8.0软件生成图表。
3 结果与分析
3.1 幼苗密度的处理间差异
5种处理的幼苗密度差异极显著(P=0.003﹤0.01)(图1),开沟处理最高(2.1株·m-2),然后依次为挖穴(1.9株·m-2)、清枯(1.8株·m-2)、割草(1.3株·m-2)和对照(1.1株·m-2)。这说明开沟、挖穴、清枯等强烈扰动的地表措施对提高幼苗密度的效果好于低度扰动的割草处理和对照。
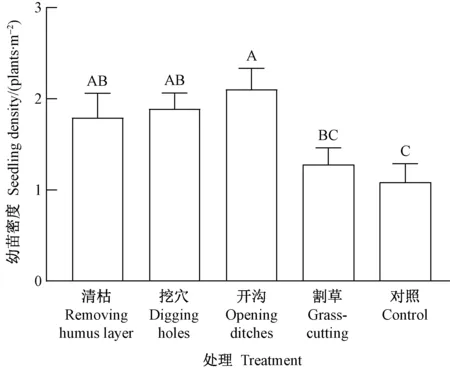
图1 幼苗密度的处理间差异
3.2 各生境类型幼苗密度的处理间差异
将5个试验点的数据依生境类型合并,比较不同处理间的幼苗密度差异。
对于林冠下生境(图2),幼苗密度的处理间差异显著(P=0.027﹤0.05),其中挖穴处理最高(2.1株,·m-2),然后依次为清枯(1.5株·m-2)、割草(1.4株·m-2)、开沟(1.3株·m-2)和对照(0.4株·m-2)。这说明林冠下挖穴处理效果最佳,调查发现幼苗主要分布在穴底四周,可能与穴壁滑落土壤覆盖种子有利于种子萌发和幼苗抗旱有关。
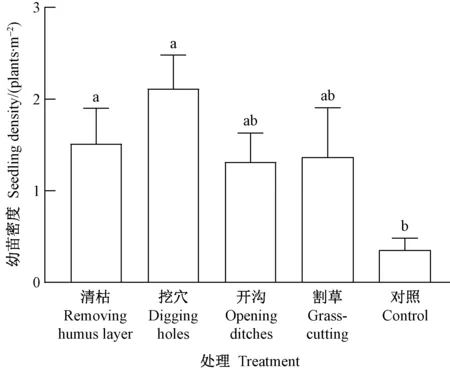
图2 林冠下生境类型幼苗密度的处理间差异
对于林隙生境(图3),幼苗密度的处理间差异极显著(P=0.003﹤0.01),其中开沟处理最高(2.4株·m-2),极显著高于其他4种处理,然后依次为挖穴(1.4株·m-2)、清枯(1.1株·m-2)、对照(1.1株·m-2)和割草(0.8株·m-2)。这说明林隙内开沟处理效果最好,可能与开沟使土壤裸露面积大、更多种子被滑落的土壤和富集的积雪覆盖有关。
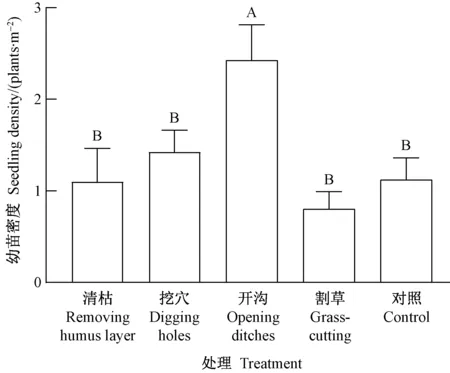
图3 林隙生境类型幼苗密度的处理间差异
对于林中空地生境(图4),幼苗密度的处理间差异不显著(P=0.276﹥0.05),其中挖穴处理最高(1.3株·m-2),然后依次为开沟(1.1株·m-2)、对照(0.8株·m-2)、清枯(0.5株·m-2)和割草(0.5株·m-2)。
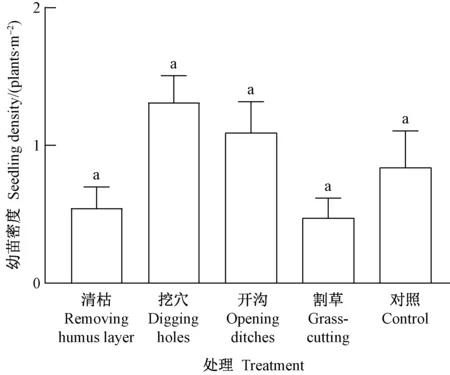
图4 林中空地生境类型幼苗密度的处理间差异
对于林缘生境(图5),幼苗密度的处理间差异极显著(P=0.000﹤0.01),其中开沟处理最高(4.1株·m-2),极显著高于其他4种处理,然后依次为割草(2.2株·m-2)、挖穴(1.2株·m-2)、清枯(0.7株·m-2)和对照(0.7株·m-2)。这说明林缘生境下开沟处理效果最好,原因与林隙生境相同。
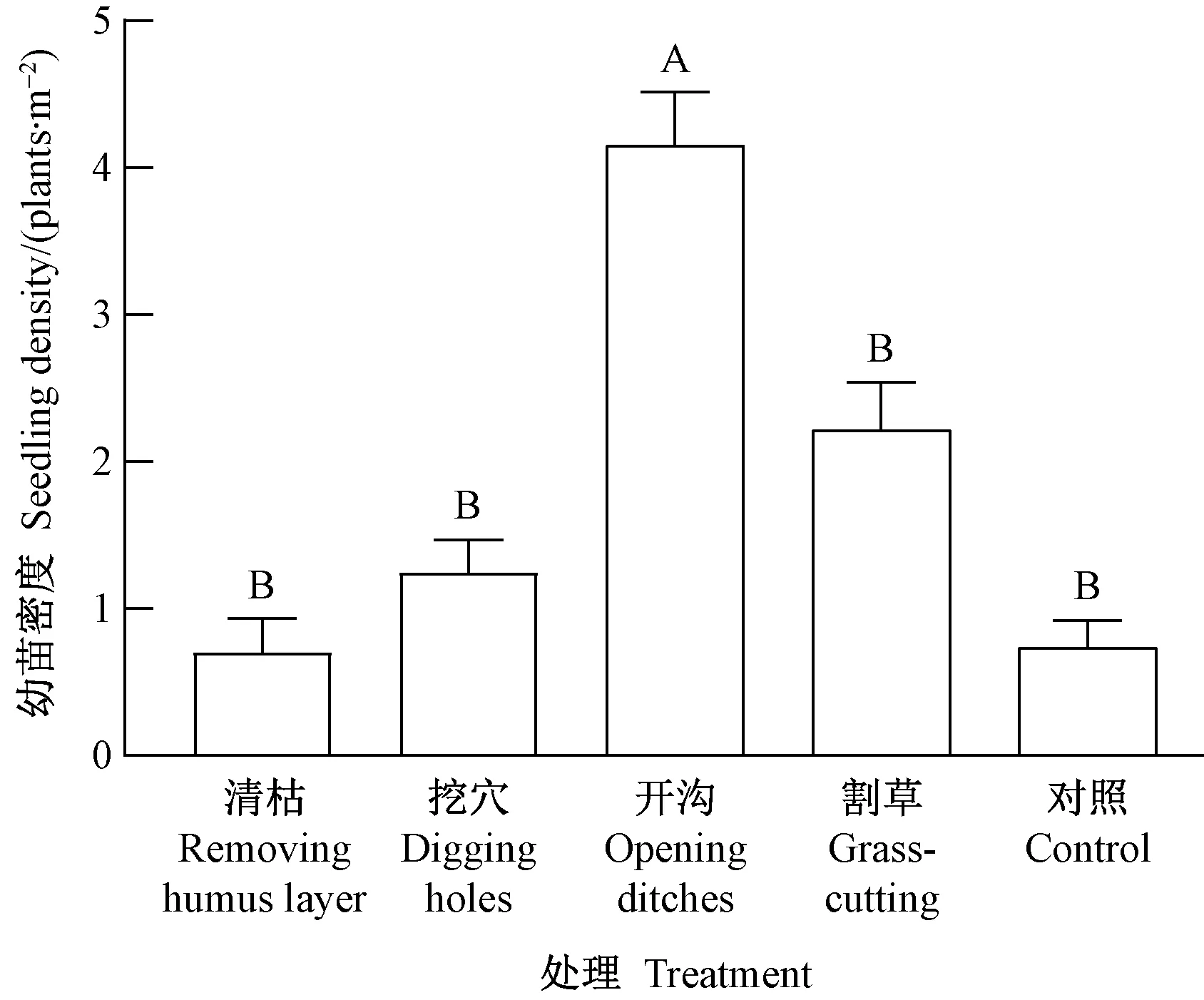
图5 林缘生境类型幼苗密度的处理间差异
对于退耕幼龄林生境(图6),幼苗密度的处理间差异不显著(P=0.380﹥0.05),其中割草处理最高(3.6),然后依次为对照(3.0株·m-2)、挖穴(2.5株·m-2)、清枯(2.4株·m-2)和开沟(2.1株·m-2)。说明在林冠稀疏、枯落物层薄的幼龄林生境,限制更新的主要因素与林冠郁闭、枯落物层厚的近熟林不同,过密杂草是重要限制因素,枯落物层限制不大,不是所有地表处理均能提高幼苗密度。

图6 退耕幼龄林生境类型幼苗密度的处理间差异
天然更新干扰因素在5类生境间的差异导致了5种处理对幼苗密度的影响不同,幼苗密度最高的处理分别是林缘的开沟(4.1株·m-2)、退耕幼龄林的割草(3.6株·m-2)、林隙的开沟2.4(株·m-2)、林冠下的挖穴2.1(株·m-2)林中空地的挖穴(1.3株·m-2)。整体来看,能去除枯落物层阻碍的开沟和挖穴、去除杂草竞争的割草和较低的林冠遮荫均有利于提高幼苗密度,但开沟效果最好。
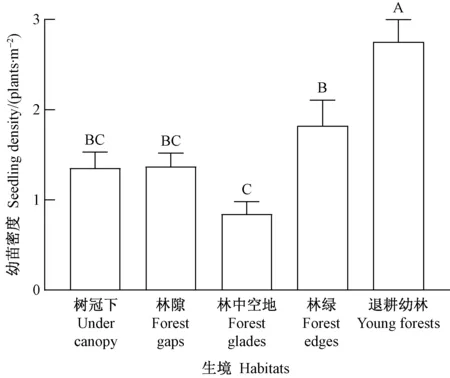
图7 不同生境类型的幼苗密度差异
3.3 各生境类型的幼苗密度差异
将5个试验点的数据合并,5类生境的幼苗密度差异极显著(P=0.000﹤0.01)(图7),其中退耕幼龄林最高(2.7株·m-2),然后依次为林缘(1.8株·m-2)、林隙(1.4株·m-2)、林冠下(1.3株·m-2)和林中空地(0.8株·m-2)。退耕幼龄林下的幼苗密度极显著高于其他生境,这与该生境林冠较稀疏(透光性合适)、枯落物层厚度不大(利于种子萌发)有关。
3.4 地面覆网对幼苗密度的影响
3.4.1 近熟林地面覆网对幼苗密度的影响 将5个试验点近熟林林冠下生境数据合并,分析覆网对幼苗密度的影响(表2)。
覆网处理的幼苗密度在5种处理间整体差异显著(P=0.049<0.05),从高到低依次为清枯(3.7株·m-2)、开沟(2.7株·m-2)、对照(1.6株·m-2)、挖穴(1.5株·m-2)和割草(0.6株·m-2),其中清枯处理效果最佳。不覆网处理幼苗密度在5种处理间整体差异显著(P=0.022﹤0.05),从高到低依次为开沟(1.7株·m-2)、清枯(1.4株·m-2)、对照(1.1株·m-2)、挖穴(1.0株·m-2)和割草(0.1株·m-2),开沟处理效果最佳。
总体分析表明,近熟林林荫下生境覆网与不覆网处理幼苗密度差异极显著(P=0.008﹤0.01),后幼荔密度均值(2.0株·m-2)极显著高于不覆网(1.1株·m-2),相差0.9株·m-2。各处理覆网均能极显著提高幼苗密度0.5~2.3株·m-2,提高幅度依次为清枯2.3株·m-2、开沟1.0株·m-2、对照0.5株·m-2、挖穴0.5株·m-2,割草0.5株·m-2。覆网可加大幼苗密度的处理间差异。
3.4.2 退耕幼龄林地面覆网对幼苗密度的影响 将5个试验点退耕幼龄林样地的数据合并,分析覆网对幼苗密度的影响(表3)。
退耕幼龄林生境幼苗密度在不同处理间的差异无论覆网(P=0.338﹥0.05)和不覆网(P=0.384﹥0.05)均不显著。覆网各处理的幼苗密度以开沟和清枯处理最高(均为4.8株·m-2),然后是挖穴(4.0株·m-2)、割草(2.7株·m-2)和对照(2.0株·m-2),最大值和最小值相差2.8株·m-2; 不覆网各处理效果排序与覆网稍有不同,依次是开沟(3.1株·m-2)、挖穴(2.7株·m-2)、清枯(2.5株·m-2)、割草(2.1株·m-2)和对照(1.7株·m-2)。表明覆网能提高幼苗密度0.3~2.3株·m-2,其中清枯物处理效果最佳。

表2 近熟林林冠下有无覆网时5种处理的幼苗密度差异①

表3 退耕幼龄林下有无覆网时5种处理的幼苗密度差异
总体分析表明,退耕幼龄林生境覆网与不覆网时幼苗密度差异显著(P=0.036﹤0.05),幼苗密度均值覆网(3.7株·m-2)比不覆网(2.4株·m-2)高1.3株·m-2。各处理覆网均能显著提高幼苗密度,提高幅度依次为清枯(2.3株·m-2)、开沟(1.7株·m-2)、挖穴(1.3株·m-2)、割草(0.6株·m-2)和对照(0.3株·m-2)。覆网可加大幼苗密度的处理间差异。
4 讨论
4.1 华北落叶松人工林能天然更新
森林天然更新是森林资源再生产中一个较为复杂的生态学过程,受多种因素影响,如种子数量和质量、立地环境(土层厚度、海拔、坡位、坡度等)、林木密度、郁闭度、草本和灌木盖度、凋落物层厚度、凋落叶中化感物质等(王笑梅等, 2017; 黄萍等, 2018; 赵总等, 2018; 黄朗等, 2019)。为了提高森林更新质量,缩短更新期,人们常采用人工更新,我国人工林现仍以人工更新为主(程中倩等, 2018),尤其是天然更新能力较弱的华北落叶松人工林。在六盘山区绝大多数华北落叶松人工林天然更新很差,几乎看不到更新幼树; 但野外调查时在一些林缘和林中空地发现有天然更新幼树,有些密度还很高,如原州区水沟林场林缘有4~5年生的华北落叶松更新幼苗,密度达21株·m-2,林中空地有5~7年生的华北落叶松更新幼苗,密度达14株·m-2。河北塞罕坝华北落叶松人工林也有大量5~120 cm高的幼苗或幼树(张树梓等, 2015)。这说明华北落叶松人工林确实能天然更新。但要充分利用天然更新,还需确定主要限制因素和作用机制,找到有效促进天然更新的技术途径,从而尽快从人工栽植更新转为天然更新或人工促进天然更新。
4.2 种子数量和活力对天然更新的影响
天然更新需要有足够多的可萌发种子。兴安落叶松(Larixgmelinii)天然更新的主要限制因素之一是种子数量(阿日根等, 2018),其林下幼苗数量与土壤及枯落物层的种子数量和活力呈显著正相关,且干旱年的种子雨数量降低也可能导致更新不佳(Ruanoetal., 2015)。但基于六盘山区华北落叶松林有较密天然更新幼树、春季和初夏常在林地看到不少幼苗等事实,本研究认为种子数量和活力不是天然更新的主要限制因素。在山西吕梁山庞泉沟,华北落叶松林种子雨密度丰年达962粒·m-2,完整种子占89%,平年达253粒·m-2(高润梅等, 2015),也表明种子数量和活力不是天然更新的最主要限制因素。
进一步说,本研究不是完全靠自然下种,而是人为撒播。张国君等(2015)研究发现,华北落叶松种子的形态、室内发芽率和1年生苗木生长指标均存在种源地的显著差异。本研究采用的种子在未催芽处理时的萌发力并不很高,室内发芽率仅31.4%,野外出苗率平均为5.5%、最高为17.0%(2017年播种出苗率),与以往报道的六盘山华北落叶松种子野外萌发率相近(杨占彪, 2009); 但水杨酸浸种可显著提高种子发芽率(范晓龙等, 2014)。按本研究撒播种子密度100粒·m-2计算(暂不考虑树木自然下种影响),假设野外出苗率平均为5.5%,则平均幼苗密度应在5.5株·m-2左右,与实测最高幼苗密度(林缘、退耕幼龄林生境的一些处理)较接近;然而多数处理的幼苗密度低于5.5株·m-2,这说明还有其他限制因素,种子数量和活力不是主要限制因素。
4.3 冬春季节种子自然损失的影响
从冬初种子落地到第2年发芽前,鼠类和鸟类会大量采食很多种子。据在山西省庞泉沟调查,4月下旬土壤种子库的完整种子只占种子库总量的60%,不完整种子占40%,其中干瘪与腐烂种子占8%,被食种子占32%,主要为鼠类啃食(高润梅等, 2015)。
本研究发现,无论是近熟林还是幼龄林的各处理,覆网均能明显提高幼苗密度,其中近熟林和幼龄林的提高幅度平均为0.9和1.3株·m-2,这表明冬春季节种子自然损失对幼苗密度的影响很大,地面覆网可有效减少动物取食,保护种子库储量(张明明, 2017)。
4.4 林地环境的影响
华北落叶松林枯落物层一般厚达几厘米,种子落地后约85%留在枯落物层,进入0~5和5~10 cm土层的仅占6%~13%和<0.5%(高润梅等, 2015)。枯落物层结构疏松,持水能力很低,连续晴天时极易干燥,因此,根系未穿透枯落物层进入水分供应相对稳定的矿质土层的大量幼苗,在其发芽、生根、生长过程中极易受干旱胁迫死亡(陈龙涛, 2017),导致成苗率很低。对山西省庞泉沟华北落叶松林的调查表明,当年6—7月出现的幼苗9月底全部死亡,并观察到死苗根茎相接处缢缩变黑,推测是水分缺失致死(高润梅等, 2015)。
在幼苗形成过程中,会遇到猝倒病、立枯病、蛴螬(Holotrichiadiomphalia)等多种病虫害,以及雨水冲刷使根系暴露的机械伤害和干旱危害,人工育苗时需采取多种防治措施(王月玲等, 2008),如用高锰酸钾或硫酸铜液浸种; 为防治猝倒病,播种时在育苗地撒施硫酸亚铁,苗木出土60%时喷洒多菌灵或退茵特液,在幼苗发病期喷洒硫酸亚铁或波尔多液; 发现立枯病时喷洒广枯灵液防治; 在苗床上灌根或喷洒辛硫磷液以防治蛴螬; 搭建遮阳网保护幼苗免受太阳暴晒和雨滴击溅等。本研究中,为模拟天然更新,未采取任何病虫害防治措施,所以病虫害导致的幼苗损失比例可能很高,未来可进行对照试验以量化病虫害的影响,并采取有效措施形成更高的幼苗密度。
幼苗生长过程中会受到林下光照及土壤水分、养分等的影响或限制。正是林地微环境的空间异质性,导致华北落叶松更新幼苗的聚集分布,且聚集程度随林分密度增加而提高(王旭刚, 2018); 华北落叶松林幼苗密度还与土壤pH、全磷和全钾含量有关(孙国龙等, 2017)。本研究中不覆网的近熟林5种处理的平均幼苗密度在林缘生境为1.8株·m-2,大于林冠下的1.3株·m-2,说明林冠过分遮荫不利于幼苗形成; 林冠郁闭度较低和枯落物层较薄的退耕幼龄林生境5种处理的平均幼苗密度为2.7株·m-2,在5类生境中最高,说明幼苗形成需要适中的郁闭度范围(董伯骞等, 2014)。可惜,本研究未能定量分析郁闭度影响并确定最佳范围,有待未来研究。
4.5 提高幼苗密度的有效措施
综上可知,限制华北落叶松天然更新的重要因素包括动物采食种子、过厚枯落物层阻碍幼苗根系进入土壤、病虫危害、过密林冠遮荫和杂草竞争等。在塞罕坝地区的研究也表明,影响天然更新幼苗密度的关键因子包括枯落物层厚度、林分密度和郁闭度等(张树梓等, 2015)。
本研究中,5类生境的平均幼苗密度表现为退耕幼龄林(2.7株·m-2)>林缘(1.8株·m-2)>林隙(1.4株·m-2)>林冠下(1.3株·m-2)>林中空地(0.8株·m-2)。这是因为在过密林中,即使暂时获得了较高幼苗密度,也难在以后存活并形成幼树(王旭刚, 2018); 间伐产生的林隙可明显促进天然更新(罗桂生等, 2019),适度间伐郁闭林分会改善更新(金红喜等, 2009)。也就是说,选择利用郁闭度较低的林分(如本研究中退耕幼龄林),制造或选择利用林隙、林窗、林缘等低遮荫生境,人工促进天然更新,会获得比过度郁闭林冠下更高的幼苗密度。
基于未覆网的所有样地调查数据,各处理的幼苗密度均值表现为开沟(2.4株·m-2)>挖穴(1.9株·m-2)=清枯(1.9株·m-2)>割草(1.4株·m-2)>对照(1.2株·m-2),与以往开沟播种可明显增加种子萌发量的研究结果(杨占彪等, 2011)相符,说明人工扰动地表有助于种子扎根到矿质土壤从而提高抗旱能力。 开沟后利于收集雨雪,开沟和挖穴后的土壤滑塌及冬季覆雪也利于保存种子,且可能因土壤滑塌形成种子覆土效果,可减少动物取食并改善水分供给; 割草处理提高幼苗密度的效果有限,是因其未彻底移除枯落物层障碍; 而彻底清除枯落物层使幼苗密度比割草和对照大幅升高。
本研究表明,地面覆网可减少动物取食从而大幅提高幼苗密度。近熟林5种处理的幼苗密度均值从不覆网的1.1株·m-2极显著提高至覆网的2.0株·m-2,平均提高0.9株·m-2,提高幅度分别为清枯2.3株·m-2、开沟1.0株·m-2、挖穴0.5株·m-2、割草0.5株·m-2、对照0.5株·m-2。退耕幼龄林5种处理的幼苗密度均值从不覆网的2.4株·m-2显著提高至覆网的3.7株·m-2,平均提高1.3株·m-2,提高幅度分别为清枯2.3株·m-2、开沟1.7株·m-2、挖穴1.3株·m-2、割草0.6株·m-2、对照0.3株·m-2。
综合来看,通过间伐形成或选择业已存在的中等郁闭生境(如未郁闭幼龄林、林缘、林隙等),在林地部分区域全面清除枯落物或大面积开沟,下种后覆盖防止动物采食种子的尼龙网,是大幅提高幼苗密度的最佳复合措施; 在林地挖穴后覆盖尼龙网的效果也较好。如在生产中应用这些措施,会利于走出华北落叶松林天然更新不足或基本没有更新的困境。然而,成功实现天然更新是一个复杂的过程,本研究仅对比了5类生境下5种处理措施对当年生幼苗夏季保留密度的影响,还未调查对当年秋末及第2年春季或更多年份后保存率的影响; 而且,从当年幼苗密度较低或很低的情况来看,未来有必要进行增大种子撒播量的梯度试验。另外,除准确确定不同条件下的林下幼苗密度和幼树生长的主要限制因素及其影响机制和数量关系外,还需定量评价不同处理措施的经济投入和投入-产出效率。这些都有待未来在更多地点开展内容更丰富、时间更长的研究。
5 结论
在宁夏六盘山地区选择华北落叶松人工林5类生境,秋末撒播种子模拟天然更新,并设置5种地表处理措施以及覆网和不覆网对照,其目的是寻找人工促进天然更新的实用技术。撒种后第2年初夏调查表明, 地面不覆网时,5类生境的平均幼苗密度从大到小依次为退耕幼龄林、林缘、林隙、林冠下和林中空地,开沟处理提高幼苗密度的效果最好; 地面覆网时,清枯处理提高幼苗密度的效果最好。限制华北落叶松天然更新的主要因素是动物取食种子、过厚枯落物层阻碍幼苗根系进入土壤、病虫危害、过密林冠遮荫和杂草竞争等,因此建议采取一些针对性措施,尤其是复合措施,如地面覆网以减少动物采食种子、清除枯落物层和扰动土壤以促进幼苗根系进入矿质土壤、选择合理的林冠郁闭生境以消除遮荫影响等,这样可大幅提高林下幼苗密度。
