IL-33/ST2信号与Treg在免疫性疾病中的研究进展*
2021-04-01李源蒋敬庭卢斌峰
李源,蒋敬庭,卢斌峰,4
(1.苏州大学附属第三医院肿瘤生物诊疗中心,江苏常州 213003;2.江苏省肿瘤免疫治疗工程技术研究中心,江苏常州 213003;3.苏州大学细胞治疗研究院,江苏常州 213003;4.美国匹兹堡大学医学院免疫学系,美国匹兹堡,PA 15213)
白细胞介素-33(interleukin-33,IL-33)基因位于人类9号染色体上,由7个外显子编码而成[1]。生长刺激表达基因2蛋白(growth stimulation expressed gene 2,ST2)基因位于染色体2q12上,长约40 000 bp。IL-33具有转录因子和细胞因子的双重功能,IL-33及其受体ST2共同参与免疫细胞之间的相互作用,在多种类型的疾病中发挥重要作用。IL-33/ST2信号可以直接作用于调节性T细胞(regulatory T cell, Treg)。Treg细胞可抑制对自身和非自身抗原的异常免疫反应,从而维持免疫稳态。本综述将探讨IL-33/ST2信号与Treg的关系及其在免疫性疾病中的研究进展,以期为多种免疫性疾病的诊疗提供思路。
1 IL-33/ST2
IL-33蛋白N末端含有一个核定位信号、一个螺旋-转角-螺旋基序样同源结构域和一个染色质结合结构域,而C端包含一个IL-1样的细胞因子结构域[1]。全长IL-33具有活性,被中性粒细胞或肥大细胞分泌的蛋白酶切割后,其活性会增强10~30倍。在凋亡过程中,IL-33被凋亡蛋白酶(胱天蛋白酶3和7)灭活,防止其激活免疫反应[2],见图1。IL-33具有转录因子和细胞因子的双重功能:全长IL-33蛋白充当核内基因调节剂,成熟的IL-33从受损或坏死细胞释放后充当细胞因子。全长IL-33以ST2不依赖的方式诱导炎症,而成熟的IL-33诱导ST2依赖的TH2相关效应。成熟IL-33通过其C端结构域与ST2受体结合,诱导构象变化,募集白介素1受体辅助蛋白(recombinant human interleukin-1 receptor accessory protein, IL-1RAcP)在细胞膜上形成异二聚体受体复合物,启动接头分子的募集,通过NF-κB、p38丝裂原活化蛋白激酶(p38 mitogen-activated protein kinase, p38MAPK)、c-Jun氨基末端激酶(c-Jun N-terminal kinase, JNK)和胞外信号调节激酶(extracellular signal-regulated kinase, EPK)激活下游转录因子。这些转录因子指导特异性细胞因子和趋化因子的表达,激活相关的下游效应[3],见图1。
ST2因与IL-33结合,被定义为IL-33受体。ST2包含一个胞外域、一个跨膜域和一个胞间域,胞外域借助IL-1RAcP结合IL-33。ST2在各种免疫细胞上均有表达,包括常规T细胞、Treg、2型先天性淋巴细胞(2 innate lymphoid cells, ILC2)、M2极化巨噬细胞、肥大细胞、嗜酸性粒细胞、嗜碱性粒细胞、嗜中性粒细胞、天然杀伤细胞(natural killer cell,NK)和iNKT细胞[3]。研究表明,IL-33/ST2信号在过敏性疾病、自身免疫病、心血管疾病等疾病中发挥重要作用[4]。患者血清中IL-33蛋白和ST2蛋白的表达水平通常采用酶联免疫吸附法检测[5-6]。
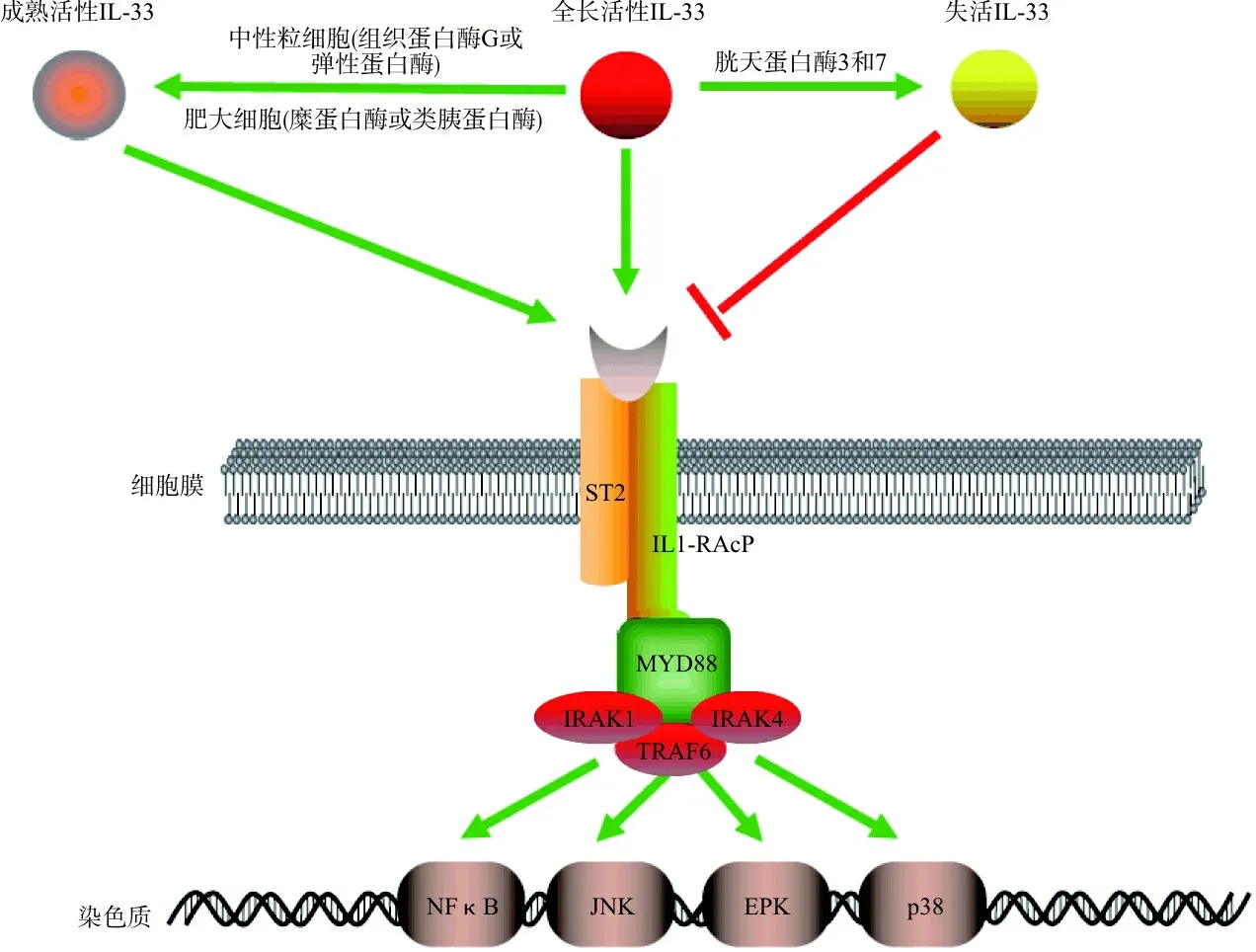
注:IL-1RAcP,白介素1受体辅助蛋白(interleukin-1 receptor accessory protein);MYD88,髓样分化因子88(myeloid differentiation factor 88);IRAK1/IRAK4,IL-1受体相关激酶1/4(interleukin-1 receptor-associated kinase1/4);TRAF6,肿瘤坏死因子受体相关因子6(TNF receptor associated factor 6);NF-κB,活化B细胞的核因子-κ轻链增强子(nuclear factor kappa-light-chain-enhancer of activated B cells);JNK, c-Jun氨基末端激酶(c-Jun N-terminal kinase);EPK,胞外信号调节激酶(extracellular signal-regulated kinase);p38, p38丝裂原活化蛋白激酶(p38 mitogen-activated protein kinase)。
2 Treg
Treg根据其起源、分化部位和表型特征不同,分为自然/胸腺来源Treg(natural source Treg, nTreg)和外周诱导Treg(peripheral induced Treg, pTreg)[7]。Treg是近年来研究较多的具有免疫抑制功能的T细胞亚群[8],通过表达Foxp3转录因子而有别于其他免疫细胞。外周血中Treg蛋白的表达通常使用流式细胞仪检测,Treg在基因水平上的表达通过RT-PCR检测Foxp3mRNA来确定[8]。虽然有几个标记可用于确定Treg细胞类型的来源,但nTreg和iTreg在体内仍很难区分[9]。研究表明,nTreg和iTreg在抗原识别上存在差异,nTreg特异性识别肿瘤细胞表面表达的自身抗原,而iTreg可能特异性识别新生抗原[10]。Foxp3+nTreg和iTreg表达CTLA4、GITR和CD25等特征性受体,这些受体进一步区别于其他免疫细胞,介导免疫抑制反应[11-12]。
3 IL-33/ST2信号与Treg
Treg发挥免疫抑制作用,可以维持免疫耐受和预防自身免疫反应[13]。研究显示,IL-33/ST2信号可将CD4+Foxp3-T细胞转化为表达Foxp3的Treg细胞[14]。此外,IL-33对于Treg细胞的生成和其抑制功能的维持必不可少[15]。因此,IL-33/ST2信号对维持Treg细胞的稳定和功能至关重要。我们将探讨IL-33/ST2信号与Treg在多种免疫性疾病中的作用。
3.1肿瘤 研究发现IL-33在肿瘤发展过程中对维持Treg细胞功能稳定具有重要作用。Foxp3+Treg细胞去除IL-33的表达导致肿瘤消退,而IL-33-/-小鼠的Treg细胞则表现出抑制作用减弱、促进肿瘤清除和增强抗肿瘤免疫[13]。IL-33通过诱导肿瘤浸润的跨膜型ST2(transmembrane ST2 isoform, ST2L)阳性Treg促进大肠癌发生发展,ST2L+Treg还可抑制CD4+CD25-T细胞的增殖和IFN-γ的产生,抑制IL-33/ST2L信号可能成为预防大肠癌的策略[16]。最近,在CT26腺癌模型中发现,对荷瘤小鼠施用重组IL-33(recombinant interleukin-33, rIL-33)促进了肿瘤组织和脾脏中ST2+Treg细胞的增殖,而阻断IL-33则可减少其增殖[16]。免疫检查点干预疗法对IL-33-/-小鼠治疗有效,αPD-1治疗可促进野生型和IL-33-/-小鼠中CD8+T细胞的扩增。此外,激活的CD8+T细胞与“脆性”Treg细胞协同作用,促进肿瘤消退,提高IL-33-/-小鼠的免疫治疗效果[13]。相反,有些研究发现IL-33/ST2信号抑制Treg细胞增殖。在黑色素瘤小鼠模型中,IL-33通过骨髓来源的抑制性细胞(myeloid-derived suppressor cells,MDSCs)降低Treg细胞分化或增殖的能力,间接抑制Treg细胞在肿瘤微环境中的浸润[17]。这些研究表明了解Treg细胞具体分子机制的重要性,以便寻找有利于抗肿瘤免疫且不会导致自身免疫的Treg治疗靶点。
3.2炎症 在T细胞诱导的结肠炎小鼠模型中,Treg细胞表达ST2对预防肠道疾病的发生至关重要。在高表达ST2+Treg细胞的肠道中,IL-33通过转化生长因子-β1介导Treg细胞分化,并为Treg细胞在炎症组织中的积累和维持提供信号[18]。此外,IL-33/ST2信号在脑Treg细胞的扩增和功能中发挥重要作用,Treg细胞通过双调蛋白(amphiregulin,Areg)抑制星形胶质细胞增生并增强神经功能,为中风和神经炎性疾病的神经元保护提供治疗机会[19]。在肺组织中也发现了同样的结果,肺Treg细胞中的IL-33信号传导诱导Areg的产生,能有效防止组织损伤[20]。在感染性疾病中,ST2+Treg细胞对炎症信号不敏感,并抑制炎症细胞因子的产生。此外,ST2+Treg细胞在小鼠感染巨细胞病毒后被诱导,在减轻肝脏损伤方面起关键作用[21]。IL-33/ST2信号在保护和促进免疫调节以及增强Treg细胞的免疫抑制功能方面发挥重要作用。然而,Treg细胞的抑制功能可能存在负面影响。研究表明,在蠕虫感染中,ST2+Treg细胞抑制TH2细胞,促进蠕虫繁殖[22]。同样,最近的一项研究显示,肿瘤释放的IL-33促进Treg细胞在促进肿瘤生长和免疫逃避的部位积累[16],具体机制仍待进一步研究。
3.3自身免疫病 Foxp3+Treg细胞在免疫稳态和维持自身耐受性方面发挥重要作用,而其缺失会导致严重的自身免疫病[23]。在自身免疫病中,IL-33在驱动或抑制T细胞反应失调中发挥重要作用。IL-33是中枢神经系统内炎症过程的主要组成部分。在自身免疫性脑炎中,IL-33抑制炎性星形胶质细胞的生成和效应T细胞的增殖,同时增强Treg和TH2反应,发挥保护作用[24]。然而,rIL-33可加重C57BL/6小鼠的自身免疫性脑炎,而阻断IL-33治疗可抑制IL-17和IFN-γ的产生[25]。这种差异可能是由于IL-33所处微环境不同,因为在疾病发作期间的IL-33信号可能是保护性的,而后期的IL-33活性可能会加剧TH1和TH17的反应[26]。IL-33与哮喘密切相关,哮喘患者IL-33的表达会增加[27],并增强气道高反应性[28]。IL-33在抗原驱动的2型气道高反应性中直接损害Treg细胞的功能[29],并通过增强与OX40配体的相互作用促进TH2的分化[30]。IL-33对Treg细胞的这种作用与之前研究不同,即IL-33损害了Treg细胞的抑制功能。上述IL-33在疾病不同时期的不同作用仍需进一步研究。
4 展望
IL-33和ST2是在免疫性疾病发展过程中协调免疫网络系统的关键信号分子。IL-33/ST2信号不仅能维持组织内稳态,还为疾病治疗提供了一个潜在靶点。作为一种细胞因子,IL-33通过与ST2结合来介导信号传导参与自身免疫和炎性疾病。IL-33/ST2信号增强TH2细胞因子的产生而获得固有和适应性免疫,并通过Treg发挥免疫抑制作用。Treg在免疫稳态和维持自身耐受性方面发挥重要作用。在一些肿瘤和自身免疫病中,IL-33的阻断药物可作为有效的治疗剂,但特定环境中IL-33的抑制机制尚不明确,仍需进行更多研究加以阐明。免疫疗法旨在调节免疫系统的活性以根除疾病。在免疫性疾病中,Treg细胞通常是进行免疫治疗的重要靶点。然而,免疫治疗在很多患者中效果欠佳,并且在应答过程中经常伴随着其他自身免疫病的发生。因此,迫切需要在不影响外周Treg细胞库的情况下精确靶向特异性Treg细胞。接下来应进一步研究IL-33/ST2信号对Treg细胞的稳定性和抑制功能的影响,了解这些疾病中Treg细胞抑制过程的分子机制,为免疫性疾病寻找新的和有效的治疗手段。
