肉鸡CMTR2基因分子特征及其表达谱分析
2020-11-02李修岭徐香慧邢晋祎
李修岭 徐香慧 邢晋祎


摘要:【目的】分析肉雞加帽甲基转移酶基因(CMTR2)分子特征及其表达谱,为后续开展鸡CMTR2基因结构及生物学功能研究打下基础。【方法】以艾拔益加(AA)肉鸡为研究对象,利用RT-PCR克隆CMTR2基因,使用DNAMAN、ProtParam、SOPMA、SWISS-MODEL、TMHMM 2.0和SignalP 3.0等在线软件进行生物信息学分析,并以实时荧光定量PCR检测分析其表达特征。【结果】肉鸡CMTR2基因编码区(CDS)序列全长为2350 bp(登录号MN296490),其开放阅读框(ORF)为2292 bp,编码763个氨基酸残基;肉鸡CMTR2氨基酸序列与原鸡CMTR2氨基酸序列的相似性为99.74%,只有2个氨基酸残基差异,与火鸡CMTR2氨基酸序列的亲缘关系较近(相似性为95.94%)。肉鸡CMTR2蛋白分子量为86855.48 Da,理论等电点(pI)为6.08,在哺乳动物网织红细胞体外表达的半衰期为30 h,在酵母体内表达的半衰期大于20 h,在大肠杆菌体内表达的半衰期大于10 h。肉鸡CMTR2蛋白为不稳定的亲水性蛋白,无信号肽和跨膜区域,包含61个磷酸化位点、15个二硫键、1个N-糖基化位点及8个O-糖基化位点;其二级结构由α-螺旋(49.93%)、无规则卷曲(35.65%)、延伸链(11.53%)和β-转角(2.88%)组成,但无法获得完整的三级结构。CMTR2基因在42日龄AA肉鸡8种组织中的相对表达量排序为脾脏>肺脏>胸肌>腹脂>肝脏>肾脏>腿肌>心脏,以在脾脏中的相对表达量最高,显著高于在其他组织中的相对表达量(P<0.05,下同);肉鸡CMTR2基因的时空表达变化趋势表现为21日龄的相对表达量最高、14日龄的相对表达量最低。【结论】CMTR2基因在不同物种中具有较高的保守性,因其在肉鸡脾脏中的表达水平最高,故推测CMTR2基因是参与机体免疫相关的关键基因。
关键词: 肉鸡;CMTR2基因;生物信息学;表达谱;机体免疫
中图分类号: S831.92 文献标志码: A 文章编号:2095-1191(2020)08-1832-08
Molecular characterization and expression pattern of
CMTR2 gene in broiler
LI Xiu-ling, XU Xiang-hui, XING Jin-yi*
(School of Life Sciences, Linyi University, Linyi, Shandong 276000, China)
Abstract:【Objective】The aim of this paper was to study the molecular characteristics and expression patterns of cap-specific mRNA (nucleoside-2'-O-)- methyltransferase 2(CMTR2) in broilers, so as to lay a foundation for further deve-lopment of CMTR2 gene structure and biological function. 【Method】RT-PCR was used to clone CMTR2 gene in Arbor Acres(AA) broilers. The bioinformatics analysis was carried out using DNAMAN, ProtParam, SOPMA, SWISS-MODEL, TMHMM 2.0 and SignalP 3.0 software, and the expression characteristics were analyzed by real-time fluorescence quantitative PCR. 【Result】The length of coding region(CDS) of CMTR2 gene in broiler was 2350 bp(accession number: MN296490), and its open reading frame(ORF) was 2292 bp, encoding 763 amino acid residues. The similarity of amino acid sequence between broiler CMTR2 and native chicken was 99.74%, only two amino acid residues were different. The phylogenetic relationship between broiler CMTR2 and Turkey CMTR2 amino acid sequence was the closest(95.94% similarity). The results showed that the molecular weight of CMTR2 protein was 86855.48 Da and the theoretical isoelectric point(PI) was 6.08. The half-life of CMTR2 protein expressed in mammalian reticulocytes in vitro was 30 h, that in yeast was more than 20 h, and that in Escherichiacoli was more than 10 h. The CMTR2 protein was an unstable hydrophilic protein with no signal peptide and transmembrane domain. It contained 61 phosphorylation sites, 15 disulfide bonds, 1 N-glycosylation site and 8 O-glycosylation sites. Its secondary structure consisted of α-helix(49.93%), random coil(35.65%), extension chain(11.53%) and β-turn(2.88%), but there was no complete tertiary structure. The relative expression of CMTR2 gene in 42-day-old AA broilers was spleen>lung>chest muscle>abdominal fat>liver>kidney>leg muscle>heart. The relative expression level of CMTR2 in spleen was the highest, which was significantly higher than that in other tissues(P<0.05, the same below); the temporal and spatial expression trend of CMTR2 gene in broilers was the highest at 21-day-old and the lowest at 14-day-old. 【Conclusion】CMTR2 gene is highly conserved in different species, and its expression level is the highest in spleen. Therefore, it is speculated that CMTR2 gene is a key gene involved in immune system.
Key words: broiler; CMTR2 gene; bioinformatics; expression patterns; body immunity
Foundation item: National Natural Science Foundation of China(31372333); Shandong Natural Science Foundation(ZR2017LC018)
0 引言
【研究意义】加帽甲基转移酶[Cap-specific mRNA (nucleoside-2'-O-)-methyltransferase 2,CMTR2]也称FTSJD1,分布在人类的细胞核和细胞质中,可催化重组RNA的核糖甲基化,对基因表达有直接影响,包括增强RNA稳定性、剪接、核质转运和翻译起始,以及促进细胞核与细胞质帽结合蛋白的相互作用等(Schroeder et al.,2004;Werner et al.,2011;Trotman and Schoenberg,2019)。鸡是重要的模式动物之一,常作为研究哺乳动物胚胎发育机制的理想材料(张月娟,2004),在生物医学研究领域发挥着重要作用,且易于饲养和便于手术操作及观察。因此,研究鸡CMTR2基因分子特征及其表达谱,可为进一步探究CMTR2基因的生物学功能积累基础资料。【前人研究进展】甲基转移酶(Methyltransferase)是动物机体内普遍存在的一种重要转移酶,能调控DNA甲基化反应,从而调节染色质结构和基因表达(苏玉等,2009;Du et al.,2015;Jensen et al.,2019)。CMTR2是一类重要的甲基转移酶,由CMTR2基因编码,其结构包括具有催化RFM(Rossmann-fold MTase)结构域的氨基端和具有非催化RFM结构域的羧基端。CMTR2的催化结构域能对mRNA帽子结构进行识别,使甲基供体与帽寡核苷酸结合形成配合物,从而促进甲基化反应(Werner et al.,2011;Smietanski et al.,2014)。人类CMTR2基因定位于16号染色体的q22.2位置上(Werner et al.,2011),而小鼠CMTR2基因定位在8号染色体的8D3位置上(Gaudet et al.,2011)。转录组测序结果表明,CMTR2基因能在多种组织中表达(Werner et al.,2011;Fagerberg et al.,2014;Yue et al.,2014);近期还在肺腺癌和鳞状细胞瘤的相关研究中发现CMTR2基因会发生显著突变,且因移码突变而富集,故推测CMTR2基因是引起此类疾病的诱因之一(Campbell et al.,2016)。此外,CMTR2基因变异与妊娠糖尿病存在显著相关性,与2型糖尿病不相关(Huerta-Chagoya et al.,2015),但CMTR2基因的生物学功能及调控机制至今尚未明确。【本研究切入点】目前,有关CMTR2基因的研究报道较少,且主要集中在酵母、病毒、小鼠及人类等方面(Saha et al.,1999;Egloff et al.,2002;Werner et al.,2011;Smietanski et al.,2014;Campbell et al.,2016),而针对鸡CMTR2基因的研究尚无报道。【拟解决的关键问题】克隆肉鸡CMTR2基因及进行生物信息学分析,并以实时荧光定量PCR检测分析其表达模式,为后续开展鸡CMTR2基因结构及生物学功能研究打下基础。
1 材料与方法
1. 1 试验材料
从山东新希望六和集团有限公司临沂种鸡场选购100羽健康状况良好的1日龄艾拔益加(AA)肉鸡,按AA肉鸡生产性能标准进行正常饲养管理和免疫接种,分别于1、7、14、21、28、35和42日齡时挑选形体大小相似的6羽AA肉鸡,禁食12 h后屠宰,采集肝脏、心脏、肺脏、腿肌、肾脏、脾脏、胸肌及腹脂等组织样品,以PBS进行冲洗,置于无菌冷冻管中,经液氮速冻后转移至-80 ℃冰箱保存备用。TRIzol(Invitrogen)和DNA凝胶回收试剂盒购自天根生化科技(北京)有限公司;M-MVL反转录酶购自Promega公司;质粒提取试剂盒购自天根生化科技(北京)有限公司;pMD19-T载体和LA Taq DNA酶购自TaKaRa公司;Hind III和EcoR I购自赛默飞世尔科技(中国)有限公司;2×SYBR Green Fast qPCR Mix购自宝日医生物技术(北京)有限公司。主要仪器设备:梯度PCR仪(Eppendorf)、冷冻离心机(Eppendorf)、凝胶成像体系(Bio-Rad)、高低温振荡培养箱(JeioTech)及超低温冰箱(Haier)等。
1. 2 总RNA提取及cDNA合成
采用TRIzol法提取各组织样品总RNA,以核酸蛋白仪(Bio-Rad)检测其OD260/OD280比值(1.8~2.1);然后采用M-MVL逆转录酶反转录合成cDNA。
1. 3 PCR引物设计及PCR扩增
根据GenBank已公布的原鸡CMTR2基因序列(XM_004944112),使用Primer Premier 6.0设计CMTR2基因扩增引物(表1)。以42日龄AA肉鸡的肝脏组织cDNA为模板,采用引物CMTR2-F和CMTR2-R进行PCR扩增。PCR反应体系25.0 μL:10×LA Buffer 2.5 μL,dNTP 2.0 μL,上、下游引物各0.4 μL,LA Taq DNA酶0.2 μL,cDNA模板1.0 μL,灭菌三蒸水18.5 μL。扩增程序:95 ℃预变性5 min;94 ℃ 30 s,59 ℃ 40 s,72 ℃ 150 s,进行35个循环;72 ℃延伸10 min。以1.2%琼脂糖凝胶电泳检测PCR扩增产物。
1. 4 肉鸡CMTR2基因测序分析
采用DNA凝胶回收试剂盒对目的条带进行纯化回收,然后连接至pMD18-T载体上,并转化大肠杆菌DH5α感受态细胞,经37 ℃过夜培养后挑取白色菌落接种至LB培养基上,摇床培养16 h,收集菌液进行PCR检测。取目标菌株提取重组质粒,以Hind III和EcoR I进行双酶切鉴定,并将阳性重组质粒送至生工生物工程(上海)股份有限公司测序。
1. 5 肉鸡CMTR2基因生物信息学分析
利用NCBI网站中的ORF Finder搜寻肉鸡CMTR2基因开放阅读框(ORF);使用DNAMAN进行肉鸡CMTR2基因核苷酸序列及其推导氨基酸序列比对分析,以MEGA 5.0中的邻接法(Neighbor-joining,NJ)构建系统发育进化树;运用ProtParam预测肉鸡CMTR2蛋白理化性质;采用SOPMA和SWISS-MODEL分别预测肉鸡CMTR2蛋白二、三级结构;使用TMHMM 2.0预测CMTR2蛋白跨膜域、PSORT II Prediction预测CMTR2蛋白亚细胞定位、SignalP 3.0预测CMTR2蛋白信号肽;并以NetPhos 3.1预测CMTR2蛋白磷酸化位点、SCRATCH Protein Prediction分析CMTR2蛋白二硫键、NetNGlyc 1.0预测CMTR2蛋白N-糖基化位点及YinOYang 1.2预测CMTR2蛋白O-糖基化位点。
1. 6 肉鸡CMTR2基因表达分析
对42日龄AA肉鸡的肝脏、心脏、肺脏、腿肌、肾脏、脾脏、胸肌及腹脂等组织进行CMTR2基因表达谱分析,以1、7、14、21、28、35和42日龄AA肉鸡肝脏组织为对照模板,分析不同發育阶段CMTR2基因的表达水平。以CMTR2-exp-F和CMTR2-exp-R为目标基因扩增引物、β-actin为内参基因(表1),采用SYBR Green I染料法进行实时荧光定量PCR检测。反应体系25.0 μL:10.0 mol/L的上、下游引物各1.0 μL,cDNA模板(10倍稀释)2.0 μL,2×SYBR Fast qPCR Mix 12.5 μL,以无菌超纯水补足至25.0 μL。扩增程序:95 ℃预变性10 min;95 ℃ 20 s,55/58 ℃ 30 s,72 ℃ 20 s,进行40个循环。每个样品进行3次重复,并通过2-△△Ct法计算CMTR2基因的相对表达量。
2 结果与分析
2. 1 肉鸡CMTR2基因克隆及测序分析结果
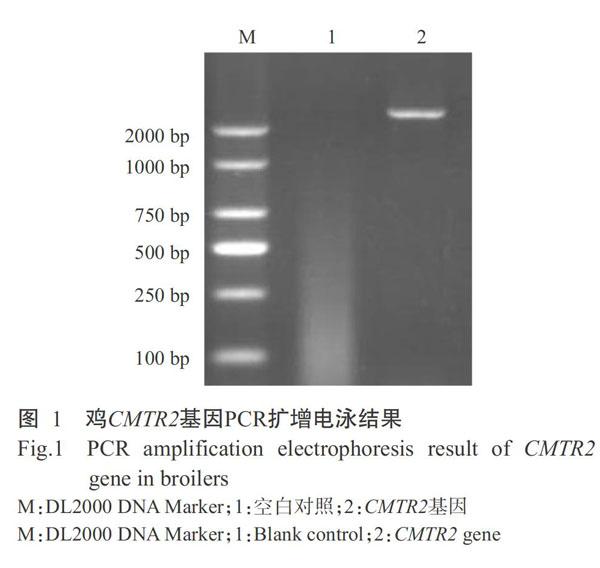
以肝脏cDNA为模板进行PCR扩增,PCR扩增产物经1.2%琼脂糖凝胶电泳检测,发现在2350 bp附近出现1条特异性条带(图1),与预期结果相符。经测序分析及序列拼接后,确认肉鸡CMTR2基因编码区(CDS)序列全长为2350 bp(登录号MN296490),其ORF为2292 bp,编码763个氨基酸残基。肉鸡CMTR2基因核苷酸序列与原鸡CMTR2基因核苷酸序列(XM_004944112)的相似性为99.66%,仅有7 bp的差异;肉鸡CMTR2氨基酸序列与原鸡CMTR2氨基酸序列的相似性为99.74%,只存在2个氨基酸残基差异,表明已成功克隆获得肉鸡CMTR2基因。
2. 2 肉鸡CMTR2氨基酸序列比对及遗传进化分析结果
由表2可知,肉鸡CMTR2氨基酸序列与火鸡CMTR2氨基酸序列的相似性最高,与鸟类CMTR2氨基酸序列的相似性均在80.00%以上,而与斑马鱼的相似性最低,为44.21%,说明CMTR2基因在物种间具有一定的保守性。基于CMTR2氨基酸序列相似性,采用邻接法构建系统发育进化树,结果显示肉鸡与火鸡和日本鹌鹑聚为一类(图2),即肉鸡与火鸡和日本鹌鹑的亲缘关系较近,与氨基酸序列比对分析结果一致。
2. 3 肉鸡CMTR2蛋白理化性质及其结构预测结果
肉鸡CMTR2蛋白分子量为86855.48 Da,理论等电点(pI)为6.08;包含82个正电荷氨基酸残基[天冬氨酸(Asp)+谷氨酸(Glu)]和67个负电荷酸残基[精氨酸(Arg)+赖氨酸(Lys)];其消光系数为93580。肉鸡CMTR2蛋白在哺乳动物网织红细胞体外表达的半衰期为30 h,在酵母体内表达的半衰期大于20 h,在大肠杆菌体内表达的半衰期大于10 h;其不稳定系数为42.33,属于不稳定蛋白。肉鸡CMTR2蛋白脂肪指数为87.77,总平均亲水性为-0.115,属于亲水性蛋白;其亚细胞定位预测结果表明主要分布在细胞质(26.1%)、分泌系统小泡(21.7%)、细胞核(13.0%)、线粒体(13.0%)、质膜(8.7%)、高尔基体(4.3%)、细胞骨架(4.3%)、空泡(4.3%)和内质网(4.3%)。
肉鸡CMTR2蛋白信号肽及跨膜结构预测结果(图3和图4)表明,不存在信号肽和跨膜区域,不属于分泌蛋白和跨膜蛋白;蛋白磷酸化预测结果显示,肉鸡CMTR2蛋白包含61个磷酸化位点(图5),分别为38个丝氨酸(Ser)磷酸化位点、18个苏氨酸(Thr)磷酸化位点和5个酪氨酸(Tyr)磷酸化位点,均大于阈值0.50,可作为蛋白激酶磷酸化位点。肉鸡CMTR2氨基酸序列含有36个半胱氨酸(Cys),即预测CMTR2蛋白中有15个二硫键。肉鸡CMTR2蛋白存在1个N-糖基化位点,位于第181位氨基酸处(图6),其潜力值为0.6668(大于阈值0.50);而在第224、305、306、311、532、671、758和760位氨基酸处有8个O-糖基化位点(图7),其潜力值分别为0.6450、0.5142、0.5131、0.5755、0.5118、0.5442、0.5606和0.5552(大于阈值0.50)。
肉鸡CMTR2蛋白二级结构由α-螺旋(49.93%)、无规则卷曲(35.65%)、延伸链(11.53%)和β-转角(2.88%)组成(图8)。肉鸡CMTR2蛋白三级结构预测结果与SWISS-MODEL数据库模板4n49只有22.48%的氨基酸序列吻合度,故无法获得其完整的三级结构,仅得到部分三级结构模型(图9)。
2. 4 肉鸡CMTR2基因表达分析结果
实时荧光定量PCR检测结果显示,CMTR2基因在42日龄AA肉鸡8种组织中的相对表达量排序为脾脏>肺脏>胸肌>腹脂>肝脏>肾脏>腿肌>心脏,以在脾脏中的相对表达量最高,显著高于在其他组织中的相对表达量(P<0.05,下同),而在心脏中的相对表达量显著低于在其他组织中的相对表达量(图10-A)。肉鸡CMTR2基因的时空表达变化趋势显示,在21日龄时的相对表达量最高,极显著高于其他日龄;在14日龄的相对表达量最低,而在28~42日龄的相对表达量呈逐渐上升趋势(图10-B)。
3 讨论
CMTR2能催化多种物质甲基化,在基因表达调控过程中发挥重要作用,进而调节动物的生长发育,但至今其具体作用机制尚不明确。本研究成功克隆获得肉鸡CMTR2基因,其CDS序列全长为2350 bp,编码763个氨基酸,与GenBank已公布的原鸡CMTR2基因核苷酸序列仅有7 bp的差异;肉鸡CMTR2氨基酸序列与原鸡CMTR2氨基酸序列仅有2个氨基酸残基差异,其余碱基突变均为同义突变。肉鸡CMTR2氨基酸序列与火鸡CMTR2氨基酸序列的相似性高达95.94%,且肉鸡、火鸡、鹌鹑、绿头鸭和鸸鹋的CMTR2氨基酸序列均为763个氨基酸残基,表明CMTR2基因在物种间高度保守。肉鸡CMTR2氨基酸序列与斑马鱼CMTR2氨基酸序列的相似性最低,说明CMTR2基因在从鱼类到鸟类经历了较强的净化选择,与系统发育进化树分析结果一致。
二硫键在蛋白分子中的主要作用是稳定其空间结构(田悦和杜军保,2007)。肉鸡CMTR2蛋白含有36个半胱氨酸(Cys)、15个二硫键,说明该蛋白抗外界因素影响的稳定性较高,且CMTR2蛋白稳定性与其半衰期成正比,即半衰期长则蛋白稳定性相应提高(贾浩等,2011)。本研究结果表明,肉鸡CMTR2蛋白在哺乳动物网状红细胞体外表达的半衰期为30 h,半衰期较长,但属于不稳定蛋白,可能与不同物种的生理功能有关。信号肽能被内质网膜上的受体识别(郑斌和詹希美,2005)。本研究的蛋白信号肽预测结果显示,肉鸡CMTR2蛋白无信号肽,说明该蛋白不属于分泌蛋白,而在胞内发挥作用,且无跨膜区域。因此,确定肉鸡CMTR2蛋白是胞内蛋白。
磷酸化修饰和糖基化修饰是机体中最常见、最重要的蛋白翻译后修饰方式,参与并调控生物机体的许多生命活动(姜铮等,2009;李军等,2009;杨妍梅等,2019)。其中,蛋白磷酸化在细胞信号转导过程中发挥重要作用(姜铮等,2009)。肉鸡CMTR2蛋白包含61个潜在的磷酸化位点,能活化CMTR2蛋白,說明该蛋白具有较高的生物活性。蛋白糖基化能增强其稳定性(李军等,2009)。肉鸡CMTR2蛋白存在1个N-糖基化位点和8个O-糖基化位点,且其二级结构以α-螺旋和无规则卷曲为主,但这种结构对CMTR2蛋白催化功能的影响还需进一步探究。由于肉鸡CMTR2蛋白三级结构仅有部分模型,且与模型中的氨基酸序列吻合度低于30.00%,故无法预测获得完整的三级结构。CMTR2蛋白三级结构主要是由α-螺旋和无规则卷曲在空间上进一步盘曲、折叠而形成,与二级结构预测结果基本一致。
本研究结果表明,肉鸡CMTR2基因在肝脏、心脏、肺脏、腿肌、肾脏、脾脏、胸肌及腹脂等8种组织中均有表达,其中,在脾脏中的相对表达量最高,在肺脏和胸肌中的相对表达量次之,与在猪(Yu et al.,2014)和大鼠(Yue et al.,2014)上的研究结果基本一致。此外,CMTR2基因在人类胸腺和睾丸(Fagerberg et al.,2014)、小鼠中枢神经系统和大脑(Yue et al.,2014)及绵羊子宫(Jiang et al.,2014)中的表达水平较高。可见,CMTR2基因对动物的免疫、繁殖、肌肉形成和生长发育均具有一定影响,但具体调控机制仍需进一步探究。
4 结论
CMTR2基因在不同物种中具有较高的保守性,因其在肉鸡脾脏中的表达水平最高,故推测CMTR2基因是参与机体免疫相关的关键基因。
参考文献:
贾浩,张小白,宋晓峰. 2011. 人类胞内蛋白半衰期与其亚细胞定位的相关性研究[J]. 计算机与应用化学,28(4):411-414. [Jia H,Zhang X B,Song X F. 2011. Relationship between intracellular proteins half-life and subcellular localization in human cells[J]. Computer and Applied Chemistry,28(4):411-414.]
姜铮,王芳,何湘,刘大伟,陈宣男,赵红庆,黄留玉,袁静. 2009. 蛋白质磷酸化修饰的研究进展[J]. 生物技术通讯,20(2):233-237. [Jiang Z,Wang F,He X,Liu D W,Chen X N,Zhao H Q,Huang L Y,Yuan J. 2009. Progress on protein/peptide phosphorylation[J]. Letters in Biotechnology,20(2):233-237.]
李军,杜鑫,Hosseini Moghaddam S H,陈玉银. 2009. 蛋白质糖基化修饰研究进展[J]. 科技通报,25(6):773-778. [Li J,Du X,Hosseini M S H,Chen Y Y. 2009. The research progress in protein glycosylation modification[J]. Bulletin of Science and Technology,25(6):773-778.]
蘇玉,王溪,朱卫国. 2009. DNA甲基转移酶的表达调控及主要生物学功能[J]. 遗传,31(11):1087-1093. [Su Y,Wang X,Zhu W G. 2009. DNA methyltransferases:The role in regulation of gene expression and biological processes[J]. Heredity (Beijing),31(11):1087-1093.]
田悦,杜军保. 2007. 二硫键和巯基在蛋白质结构功能中的作用及分析方法[J]. 实用儿科临床杂志,22(19):1499-1501. [Tian Y,Du J B. 2007. Effect of disulfide bond and mercap to group on structure and function of protein and analytical method[J]. Chinese Journal of Applied Clinical Pediatrics,22(19):1499-1501.]
杨妍梅,李玉,覃圣,蒋刘芽,赵金潭,陆晓东,熊杰,陈伟基,扎西英派. 2019. 静宁鸡PPARα基因克隆与生物信息学分析[J]. 江苏农业学报,35(2):370-377. [Yang Y M,Li Y,Qin S,Jiang L Y,Zhao J T,Lu X D,Xiong J,Chen W J,Zhaxi Y P. 2019. Cloning and bioinformatic analysis of PPARα gene in Jingning chicken[J]. Jiangsu Journal of Agricultural Sciences,35(2):370-377.]
张月娟. 2004. 模式动物鸡和果蝇的心脏发育研究[D]. 长沙:湖南师范大学. [Zhang Y J. 2004. Research of the heart development in chick and Drosophila[D]. Changsha:Hunan Normal University.]
郑斌,詹希美. 2005. 信号肽序列及其在蛋白质表达中的应用[J]. 生物技术通讯,16(3):296-298. [Zheng B,Zhan X M. 2005. Signal peptide sequence and its application in protein expression[J]. Letters in Biotechnology,16(3):296-298.]
Campbell J D,Alexandrov A,Kim J,Wala J,Berger A H,Peda-mallu C S,Shukla S A,Guo G W,Brooks A N,Murray B A,Imielinski M,Hu X,Ling S Y,Akbani R,Rosenberg M,Cibulskis C,Ramachandran A,Collisson E A,Kwiatkowski D J,Lawrence M S,Weinstein J N,Verhaak R G,Wu C J,Hammerman P S,Cherniack A D,Getz G,Cancer Genome Atlas Research Network,Artyomov M N,Schreiber R,Govindan R,Meyerson M. 2016. Distinct patterns of somatic genome alterations in lung adenocarcinomas and squamous cell carcinomas[J]. Nature Genetics,48(6):607-616.
Du J M,Johnson L M,Jacobsen S E,Patel D J. 2015. DNA methylation pathways and their crosstalk with histone methylation[J]. Nature Reviews. Molecular Cell Biology,16(9):519-532.
Egloff M P,Benarroch D,Selisko B,Romette J L,Canard B. 2002. An RNA cap(nucleoside-2'-O-)-methyltransferase in the flavivirus RNA polymerase NS5:Crystal structure and functional characterization[J]. The EMBO Journal,21(11):2757-2768.
Fagerberg L,Hallstr?m B M,Oksvold P,Kampf C,Djurei-novic D,Odeberg J,Habuka M,Tahmasebpoor S,Da-nielsson A,Edlund K,Asplund A,Sj?stedt E,Lundberg E,Szigyarto C A K,Skogs M,Takanen J O,Berling H,Tegel H,Mulder J,Nilsson P,Schwenk J M,Lindskog C,Danielsson F,Mardinoglu A,Sivertsson A,von Feilitzen K,Forsberg M,Zwahlen M,Olsson I,Navani S,Huss M,Nielsen J,Ponten F,Uhlén M. 2014. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics[J]. Molecular & Cellular Proteomics,13(2):397-406.
Gaudet P,Livstone M S,Lewis S E,Thomas P D. 2011. Phylogenetic-based propagation of functional annotations within the gene ontology consortium[J]. Briefings in Bio-informatics,12(5):449-462.
Huerta-Chagoya A,Vázquez-Cárdenas P,Moreno-Macías H,Tapia-Maruri L,Rodríguez-Guillén R,López-Vite E,García-Escalante G,Escobedo-Aguirre F,Parra-Covarrubias A,Cordero-Brie?o R,Manzo-Carrillo L,Zacarías-Castillo R,Vargas-García C,Aguilar-Salinas C,Tusié-Luna T. 2015. Genetic determinants for gestational diabetes mellitus and related metabolic traits in Mexican women[J]. PLoS One,10(5):e0126408.
Jensen T ?,Tellgren-Roth C,Redl S,Maury J,Jacobsen S A B,Pedersen L E,Nielsen A T. 2019. Genome-wide systematic identification of methyltransferase recognition and modification patterns[J]. Nature Communications,10(1):3311.
Jiang Y,Xie M,Chen W B,Talbot R,Maddox J F,Faraut T,Wu C H,Muzny D M,Li Y X,Zhang W G,Stanton J A,Brauning R,Barris W C,Hourlier T,Aken B L,Searle S M J,Adelson D L,Bian C,Cam G R,Chen Y L,Cheng S F,DeSilva U,Dixen K,Dong Y,Fan G Y,Franklin I R,Fu S Y,Guan R,Highland M A,Holder M E,Huang G D,Ingham A B,Jhangiani S N,Kalra D,Kovar C L,Lee S L,Liu W Q,Liu X,Lu C X,Lü T,Mathew T,McWilliam S,Menzies M,Pan S K,Robelin D,Servin B,Townley D,Wang W L,Wei B,White S N,Yang X H,Ye C,Yue Y J,Zeng P,Zhou Q,Hansen J B,Kristensen K,Gibbs R A,Flicek P,Warkup C C,Jones H E,Oddy V H,Nicholas F W,McEwan J C,Kijas J,Wang J,Worley K C,Archibald A L,Cockett N,Xu X,Wang W,Dalrymple B P. 2014. The sheep genome illuminates biology of the rumen and lipid metabolism[J]. Science,344(6188):1168-1173.
Saha N,Schwer B,Shuman S. 1999. Characterization of human,Schizosaccharomyces pombe,and Candida albicans mRNA cap methyltransferases and complete replacement of the yeast capping apparatus by mammalian enzymes[J]. The Journal of Biological Chemistry,274(23):16553-16562.
Schroeder S C,Zorio D A R,Schwer B,Shuman S,Bentley D. 2004. A function of yeast mRNA cap methyltransfera-se,Abd1,in transcription by RNA polymerase II[J]. Molecular Cell,13(3):377-387.
Smietanski M,Werner M,Purta E,Kaminska K H,Stepinski J,Darzynkiewicz E,Nowotny M,Bujnicki J M. 2014. Structural analysis of human 2'-O-ribose methyltransfera-ses involved in mRNA cap structure formation[J]. Nature Communications,5:3004. doi:10.1038/ncomms4004.
Trotman J B,Schoenberg D R. 2019. A recap of RNA recapping[J]. Wiley Interdisciplinary Reviews. RNA,10(1):e1504.
Werner M,Purta E,Kaminska K H,Cymerman I A,Campbell D A,Mittra B,Zamudio J R,Sturm N R,Jaworski J,Bujnicki J M. 2011. 2'-O-ribose methylation of cap2 in human:Function and evolution in a horizontally mobile family[J]. Nucleic Acids Research,39(11):4756-4768.
Yu Y,Fuscoe J C,Zhao C,Guo C,Jia M W,Qing T,Bannon D I,Lancashire L,Bao W J,Du T T,Luo H,Su Z Q,Jones W D,Moland C L,Branham W S,Qian F,Ning B T,Li Y,Hong H X,Guo L,Mei N,Shi T L,Wang K Y,Wolfinger R D,Nikolsky Y,Walker S J,Duerksen-Hughes P,Mason C E,Tong W D,Thierry-Mieg J,Thierry-Mieg D,Shi L M,Wang C. 2014. A rat RNA-Seq transcriptomic BodyMap across 11 organs and 4 developmental stages[J]. Nature Communications,5:3230. doi:10. 1038/ncomms4230.
Yue F,Cheng Y,Breschi A,Vierstra J,Wu W S,Ryba T,Sandstrom R,Ma Z H,Davis C,Pope B D,Shen Y,Pervouchine D D,Djebali S,Thurman R E,Kaul R,Rynes E,Kirilusha A,Marinov G K,Williams B A,Trout D,Amrhein H,Fisher-Aylor K,Antoshechkin I,DeSalvo G,See L H,Fastuca M,Drenkow J,Zaleski C,Dobin A,Prie-to P,Lagarde J,Bussotti G,Tanzer A,Denas O,Li K,Bender M A,Zhang M H,Byron R,Groudine M T,McCleary D,Pham L,Ye Z,Kuan S,Edsall L,Wu Y C,Rasmussen M D,Bansal M S,Kellis M,Keller C A,Morrissey C S,Mishra T,Jain D,Dogan N,Harris R S,Cayting P,Kawli T,Boyle A P,Euskirchen G,Kundaje A,Lin S,Lin Y,Jansen C,Malladi V S,Cline M S,Erickson D T,Kirkup V M,Learned K,Sloan C A,Rosenbloom K R,de Sousa B L,Beal K,Pignatelli M,Flicek P,Lian J,Kahveci T,Lee D,Kent W J,Santos M R,Herrero J,Notredame C,Johnson A,Vong S,Lee K,Bates D,Neri F,Diegel M,Canfield T,Sabo P J,Wilken M S,Reh T A,Giste E,Shafer A,Kutyavin T,Haugen E,Dunn D,Reynolds A P,Neph S,Humbert R,Hansen R S,de Bruijn M,Selleri L,Rudensky A,Josefowicz S,Samstein R,Eichler E E,Orkin S H,Levasseur D,Papayannopoulou T,Chang K H,Skoultchi A,Gosh S,Disteche C,Treuting P,Wang Y L,Weiss M J,Blobel G A,Cao X Y,Zhong S,Wang T,Good P J,Lowdon R F,Adams L B,Zhou X Q,Pazin M J,Feingold E A,Wold B,Taylor J,Mortazavi A,Weissman S M,Stamatoyannopoulos J A,Snyder M P,Guigo R,Gingeras T R,Gilbert D M,Hardison R C,Beer M A,Ren B. 2014. A comparative encyclopedia of DNA elements in the mouse genome[J]. Nature,515(7527):355-364.
(責任编辑 兰宗宝)
收稿日期: 2019-09-07
基金项目:国家自然科学基金项目(31372333);山东省自然科学基金项目(ZR2017LC018)
作者简介:*为通讯作者,邢晋祎(1968-),博士,教授,主要从事动物遗传育种研究工作,E-mail:xingjinyi@lyu.edu.cn。李修岭(1972-),教授,主要从事动物分子遗传进化研究工作,E-mail:379124952@qq.com
