miR-30c-2-3p靶向MPO介导高糖诱导的肾小球系膜细胞凋亡及炎性反应
2020-09-09张娟陈家斌陈龙彭沛余凤荣
张娟 陈家斌 陈龙 彭沛 余凤荣
糖尿病肾病(DN)是导致糖尿病及慢性肾衰竭死亡的重要原因之一,主要表现为肾小球系膜细胞增生小管间质扩张,肾小球系膜区和小管间质细胞外基质积聚等,最终导致肾小球硬化、肾脏纤维化[1]。DN的发病机制尚未完全阐明,研究发现多种miRNA参与系膜细胞损伤,在肾脏纤维化过程中发挥重要作用[2];还参与肾小管间质纤维化,与糖尿病肾病的发生、发展相关[3]。缺氧诱导因子HIF2α在透明细胞肾细胞癌中能增强肿瘤生长,而抑制miR-30c-2-3p表达可以增强HIF2α表达,说明miR-30c-2-3p与肾细胞癌的进展相关[4]。增加miR-30c-2-3p的表达可提高足细胞的活力,同时可增加Nephrin和Podocin蛋白的表达[5]。但miR-30c-2-3p对肾小球系膜细胞凋亡的影响还不清楚。髓过氧化物酶(myeloperoxidase,MPO)是血红素过氧化物酶超家族成员之一,也是一种炎性指标,在炎症的发生发展过程中具有重要作用,与多种疾病相关[6]。在高糖的人肾小球系膜细胞中MPO、蛋白的表达增加[7],MPO与空腹血糖水平呈正相关,在糖尿病微血管病变机制中起着十分重要的作用,检测MPO及空腹血糖浓度,对于糖尿病并发微血管病变的预防和治疗具有重要的意义[8]。但miR-30c-2-3p是否通过MPO影响肾小球系膜细胞的进展还不清楚,本文旨在研究miR-30c-2-3p对高糖诱导的肾小球系膜细胞凋亡及炎症反应的影响以及是否通过MPO发挥作用,为糖尿病肾病的诊断、治疗和预后提供新的思路和新靶点。
1 材料与方法
1.1 材料 人肾小球系膜细胞HMC购自中国科学院上海细胞库;胎牛血清、DMEM培养基、胰蛋白酶、二甲基亚砜(DMSO)、BCA试剂盒、PBS缓冲液购自美国Sigma公司;Lipofectamine TM 2000转染试剂盒购自美国Invitrogen公司;Trizol试剂、反转录试剂盒、荧光定量试剂盒购自日本TaKaRa公司;膜联蛋白V-异硫氰酸荧光素(Annexin V-FITC)和碘化丙锭(PI)试剂盒、双荧光素酶报告基因检测试剂盒购自北京Solarbio公司;抗体均购自上海煊翎生物科技有限公司;酶联免疫吸附试验(ELISA)检测试剂盒购自上海联科生物技术有限公司;RIPA蛋白裂解液、PVDF膜、SDS-PAGE试剂盒购自上海碧云天生物技术有限公司。
1.2 方法
1.2.1 细胞培养与分组:将肾小球系膜细胞HMC在37℃、含10%胎牛血清的DMEM完全培养液中培养,每天换液一次,将融和至80%左右的细胞无血清饥饿培养24 h后进行分组,用终浓度为30 mmol/L的葡萄糖溶液刺激细胞作为高糖组(HG),培养48 h后收集细胞,同时以终浓度为5 mmol/L的葡萄糖溶液处理细胞作为正常对照组(NG)。将miR-NC,miR-30c-2-3p、anti-miR-NC、anti-miR-30c-2-3p分别转染至未经任何处理的HMC细胞中,记为miR-NC组,miR-30c-2-3p组、anti-miR-NC组、anti-miR-30c-2-3p组。将miR-NC、miR-30c-2-3p转染至高糖处理的HMC细胞中,记为HG+miR-NC组、HG+miR-30c-2-3p组。将miR-30c-2-3p与pcDNA3.1、pcDNA3.1-MPO分别共同转染至高糖处理的HMC细胞中,记为HG+miR-30c-2-3p+pcDNA3.1组、HG+miR-30c-2-3p+pcDNA3.1-MPO组;转染均采用Lipofectamine TM 2000转染试剂盒。
1.2.2 qRT-PCR检测miR-30c-2-3p的表达水平:收集各组细胞,研磨充分后加入Trizol试剂提取总RNA,微量核酸测定仪检测RNA纯度和浓度。使用TaKaRa反转录试剂盒将RNA反转录成cDNA,按照TaKaRa 荧光定量试剂盒使用说明配制反应体系,以β-actin为内参进行PCR扩增,每个样品重复3次,循环条件为95℃ 30 s,60℃ 30 s;72℃ 30 s,共40个循环;72℃延长5 min。相对表达量采用2-ΔΔCt法计算。
1.2.3 Western blot检测蛋白表达:收集各组细胞,加入RIPA裂解液裂解,4℃,12 000 g离心15 min,收集蛋白上清液,BCA试剂盒测定蛋白浓度。将蛋白样品进行SDS-PAGE电泳后转至PVDF膜上,5%脱脂奶粉封闭液室温封闭1 h。分别加入一抗(1∶1 000),4℃孵育过夜,TBST洗膜;加入二抗(1∶2 000)室温孵育2 h,TBST洗涤3次,每次10 min,后在暗室中曝光显影,再浸入定影,最后洗去残液晾干,将胶片用Quantity One凝胶分析软件处理,测定各组蛋白条带的吸光度,以目的条带和GAPDH条带的比值作为蛋白表达水平。每个蛋白样品重复3次。
1.2.4 流式细胞术检测细胞凋亡:用不含EDTA的胰酶消化各组细胞,离心收集各组细胞,PBS漂洗2次,加结合缓冲液重悬细胞。依据试剂盒说明书,先后加入Annexin V-FITC和PI避光孵育15 min。流式细胞仪检测激发波长488 nm和发射波长530 nm处的荧光强度。实验重复3次。
1.2.5 ELISA法检测TNF-α和IL-6的表达:取各组细胞,离心取上清,然后按照ELISA试剂盒说明书进行检测。
1.2.6 荧光素酶报告基因检测实验检测miR-30c-2-3p对MPO的靶向调控:TargetScan数据库显示MPO 3′UTR区域有miR-30c-2-3p结合位点。构建野生型和突变型基因靶点MPO的3′UTR-荧光素酶表达载体(WT-MPO和MUT-MPO),取对数生长期小鼠足细胞接种于24孔板(5×104个/孔),待细胞生长至80%融合时,用LipofectamineTM 2000将WT-MPO和MUT-MPO组细胞分别转染miR-NC和miR-30c-2-3p。依据说明书要求,使用荧光素酶报告基因检测仪进行双荧光素酶报告实验测定。实验结果以荧光素酶活性和Renilla活性的比值进行统计学分析。实验重复3次。
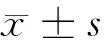
2 结果
2.1 miR-30c-2-3p在高糖作用的细胞HMC中的表达 qRT-PCR检测结果显示,与NG组相比,HG组细胞HMC中miR-30c-2-3p的表达水平显著降低(P<0.05)。可见,miR-30c-2-3p在高糖作用的细胞HMC中低表达。见表1。

表1 miR-30c-2-3p在高糖作用的细胞HMC中的表达
2.2 过表达miR-30c-2-3p对高糖作用的细胞HMC凋亡的影响 qRT-PCR检测结果显示,与HG+miR-NC组相比,HG+miR-30c-2-3p组HMC细胞中miR-30c-2-3p的表达水平显著升高(P<0.05)。Western Blot检测结果显示,与HG+miR-NC组相比,HG+miR-30c-2-3p组HMC细胞中Bcl-2蛋白的表达水平显著升高,Bax蛋白的表达水平显著降低(P<0.05)。流式细胞仪检测结果显示,与HG+miR-NC组相比,HG+miR-30c-2-3p组HMC细胞的凋亡率显著降低(P<0.05)。可见,过表达miR-30c-2-3p可抑制高糖作用的HMC细胞凋亡。见图1,表2。

图1 过表达miR-30c-2-3p对高糖作用的细胞HMC凋亡的影响;A 过表达miR-30c-2-3p对高糖作用的细胞HMC凋亡的影响;B 凋亡相关蛋白的表达
表2 过表达miR-30c-2-3p对高糖作用的细胞HMC凋亡的影响
2.3 过表达miR-30c-2-3p对高糖作用的细胞HMC中IL-6、TNF-α表达的影响 ELISA法检测结果显示,与HG+miR-NC组相比,HG+miR-30c-2-3p组HMC细胞中IL-6和TNF-α的表达水平显著降低(P<0.05)。可见,过表达miR-30c-2-3p抑制高糖作用的细胞HMC中IL-6、TNF-α的表达。见表3。
表3 过表达miR-30c-2-3p对高糖作用的细胞HMC中IL-6、TNF-α表达的影响
2.4 miR-30c-2-3p靶向、调控MPO 通过TargetScan数据库预测到MPO与miR-30c-2-3p存在结合位点。荧光素酶报告基因检测实验结果显示,转染野生型MPO基因表达载体WT-MPO后,相较于miR-NC组,miR-30c-2-3p组WT-MPO肾小球系膜细胞的荧光素酶活性显著降低(P<0.05);而转染突变型MPO基因表达载体MUT-MPO后,相较于miR-NC组,miR-30c-2-3p组MUT-MPO肾小球系膜细胞的荧光素酶活性差异不显著。Western Blot检测结果显示,相较于miR-NC组,miR-30c-2-3p组肾小球系膜细胞中MPO蛋白的表达水平显著降低;相较于anti-miR-NC组,anti-miR-30c-2-3p组肾小球系膜细胞中MPO蛋白的表达水平显著升高(P<0.05)。可见,miR-99a-3p可靶向调控MPO的表达。见图2,表4、5。
表4 双荧光素酶报告实验
表5 miR-30c-2-3p调控MPO的表达
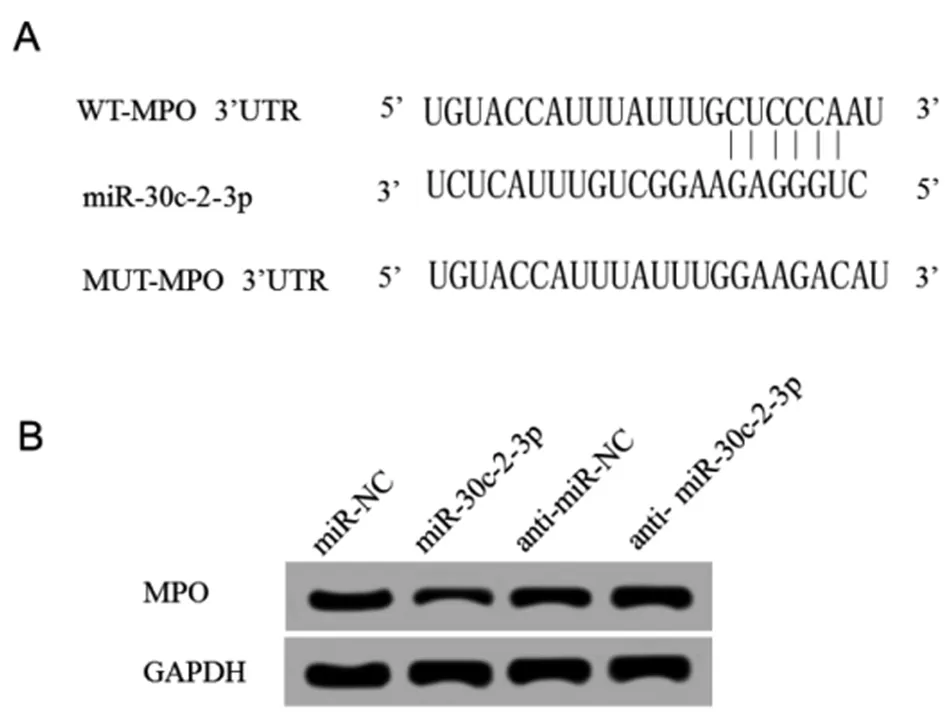
图2 miR-30c-2-3p靶向、调控MPOA:MPO的3’UTR含有miR-30c-2-3p的互补序列 B:miR-30c-2-3p调控MPO的表达
2.5 过表达MPO能逆转miR-30c-3p对高糖作用的细胞HMC凋亡及IL-6、TNF-α表达的影响 Western Blot检测结果显示,与HG+miR-NC组相比,HG+miR-30c-2-3p组HMC细胞中Bcl-2蛋白的表达水平显著升高,Bax、MPO蛋白的表达水平显著降低;与HG+miR-30c-2-3p+pcDNA3.1组相比,HG+miR-30c-2-3p+pcDNA3.1-MPO组HMC细胞中Bcl-2蛋白的表达水平显著降低,Bax、MPO蛋白的表达水平显著升高(P<0.05)。流式细胞仪检测结果显示,与HG+miR-NC组相比,HG+miR-30c-2-3p组HMC细胞的凋亡率显著降低;与HG+miR-30c-2-3p+pcDNA3.1组相比,HG+miR-30c-2-3p+pcDNA3.1-MPO组HMC细胞的凋亡率显著升高(P<0.05)。ELISA法检测结果显示,与HG+miR-NC组相比,HG+miR-30c-2-3p组HMC细胞中IL-6和TNF-α的表达水平显著降低;与HG+miR-30c-2-3p+pcDNA3.1组相比,HG+miR-30c-2-3p+pcDNA3.1-MPO组HMC细胞中IL-6和TNF-α的表达水平显著升高(P<0.05)。可见,过表达MPO能逆转miR-30c-3p对高糖作用的细胞HMC凋亡及IL-6、TNF-α表达的抑制作用。见图3,表6、7。

图3 过表达MPO能逆转miR-30c-3p对高糖作用的细胞HMC凋亡蛋白表达的影响
表6 过表达MPO能逆转miR-30c-3p对高糖作用的细胞HMC凋亡的影响
表7 过表达MPO能逆转miR-30c-3p对高糖作用的细胞中IL-6、TNF-α表达的影响
3 讨论
DN最终导致的终末期肾功能衰竭是糖尿病患者死亡率高的原因之一,严重威胁着人类的健康,肾小球系膜细胞的异常增殖是早期DN的重要特征,研究其具体的作用机制有助于早期诊断和预防[9]。研究发现许多miRNAs在肾脏发育、进展和相关疾病中扮演重要角色[10],且在DN中异常表达,参与DN的发病机制[11]。研究发现miR-30c-2-3p在大鼠心肌细胞缺氧/复氧损伤模型中高表达,且抑制miR-30c-2-3p可通过上调XBP1表达减轻心肌细胞缺氧/复氧损伤[12]。miR-30c-2-3p还通过下调乳腺癌中的TRADD和CCNE1负调节NF-κB信号传导和细胞周期进程[13];有研究通过对原发性局灶节段硬化性肾小球肾炎患者血浆和正常人血浆进行miRNA芯片分析,发现miR-30c-2-3p在肾炎患者中低表达[14]。本实验结果显示,miR-30c-2-3p在高糖诱导的肾小球系膜细胞中低表达,过表达miR-30c-2-3p表达可抑制高糖作用的HMC细胞凋亡,抑制IL-6、TNF-α的表达。
MPO主要是由中性粒细胞和单核细胞分泌的一种过氧化物酶,作为一种新型炎症细胞因子与机体免疫有关,可能通过炎症反应和免疫应答等诱发疾病的发生[15]。MPO可能通过急性期炎性反应和氧化应激参与终末期肾病的病理过程[16]。研究发现MPO抑制体外培养的内皮细胞的生长和增殖,促进内皮细胞凋亡[17];MPO和IL-6在急性心肌梗死患者中高表达,可作为急性心肌梗死的早期诊断标记物[18]。高糖能使肾小球系膜细胞上清液中MPO活性增加,导致人肾小球系膜细胞氧化失衡[19]。抗MPO抗体可作为肾炎临床鉴别诊断的参考指标[20]。IL-6、TNF-α也是炎症相关因子,在糖尿病患者中含量较高,TNF-α可反映糖尿病患者肾脏损伤程度[21]。本实验结果显示,miR-30c-2-3p靶向调控MPO的表达,过表达MPO能逆转miR-30c-2-3p过表达对高糖作用的HMC细胞凋亡及IL-6、TNF-α表达的抑制作用。
综上所述,miR-30c-2-3p能抑制高糖作用的HMC细胞凋亡,其机制主要是下调MPO及炎性因子IL-6、TNF-α的表达。可为糖尿病肾病的治疗提供新思路和新靶点。
