上海市伪狂犬病病毒相关毒力基因分子变异分析
2020-07-06
(上海市动物疫病预防控制中心,上海 201103)
伪狂犬病病 毒(pseudorabies virus,PRV)又名猪疱疹病毒I 型(porcine herpesvirus type I),属于疱疹病毒科(Herpesviridae)甲亚科(Alphaerpesvirinae)水痘病毒属(Varicellovirus),为线状双股DNA 病毒,全长约150 kb,含77个读码框,平均G+C 含量高达73.6%。PRV 由独特的长区段(unique long region,UL)和短区段(unique short region,US)及位于US 两侧的末端重复序列(TR)和内部重复序列(IR)所组成,即形成了UL-IR-US-IR 结构[1]。
PRV 毒力由多基因控制。与其毒力相关的蛋白可分为3 类:囊膜糖蛋白、病毒自身的酶和非必需的衣壳蛋白。糖蛋白gB 是PRV 的必需囊膜糖蛋白,位于UL27 区,全长2 745 bp,编码914个氨基酸。gB 是重要的免疫原,能够刺激机体产生补体依赖性和非依赖性中和抗体[2]。gB 在病毒增殖和感染过程中参与细胞融合过程,是病毒复制必不可少的[3]。糖蛋白gC 是PRV 的重要囊膜蛋白,位于UL44 区,全长1 464 bp,编码488个氨基酸。尽管gC 对病毒感染是非必需的,但它可刺激机体产生补体非依赖性中和抗体,还能诱导细胞免疫[5-6]。gB、gC 糖蛋白是PRV 的主要T 细胞抗原,也是诱导体液免疫的重要中和抗原,与病毒释放密切相关[7]。gD 蛋白是PRV 的必需糖蛋白,位于US6 区,全长1 209 bp,编码402个氨基酸,具有糖蛋白固有的特征。gD 能识别细胞表面的受体,介导病毒与靶细胞的稳定结合,同时能够诱导机体产生中和抗体。gE 蛋白是PRV 的一种非必需糖蛋白,位于US 区,长1 734 bp,编码577个氨基酸,具有典型的膜蛋白特征。gE是PRV 主要的毒力基因,能介导细胞的融合,并有助于PRV 向中枢神经系统扩散。
本研究利用PCR 技术,对2010—2012年分离的28 株PRV 毒株的4种相关毒力基因(gB、gC、gD和gE)进行克隆、测序和相关生物学分析,旨在了解上海市PRV 主要毒力基因的分子特征及变异情况,为PRV 分子致病机制及分子流行病学研究提供依据。
1 材料与方法
1.1 毒株
28 株PRV 由本中心实验室于2010—2015年分离鉴定保存,具体信息见表1。
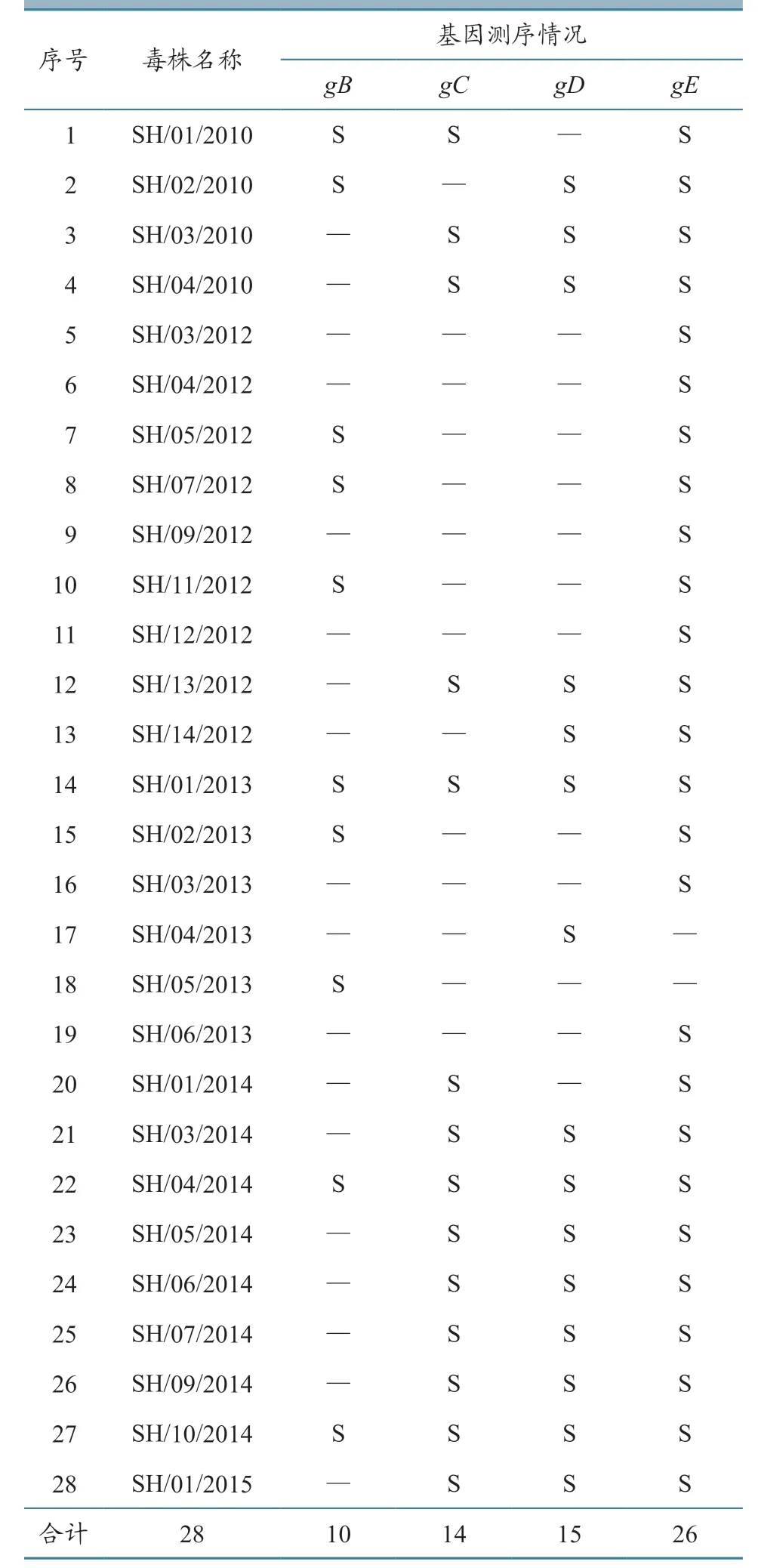
表1 本研究所用毒株
1.2 主要试剂
核酸提取试剂盒QIAamp MinElute Virus Spin Kit,购 自QIAGEN 公 司;PrimeSTAR HS DNA Polymerse with GC Buffer、DL 2 000 Marker、DNA胶回收试剂盒和pMD 18-T 载体,均购自宝生物工程(大连)有限公司。
1.3 引物
根据PRVgB、gC、gD和gE基因核苷酸保守序列,采用Primer 5.0 软件设计引物(表2)。引物由上海Invitrogen 公司或上海桑尼生物科技有限公司合成。

表2 本研究所用引物
1.4 核酸提取及PCR
参照QIAGEN 公司病毒核酸提取试剂盒(QIAamp MinElute Virus Spin Kit)说明提取病毒核酸。
PCR 反应体系 为50 µL:2×PrimeSTAR GC Buffer(Mg2+plus)25 µL,上下游引物各1 µL(20 pmol/L),dNTP Mixture 2 µL,上 述DNA 5 µL,补H2O 至50 µL。PCR 反应条件为:95 ℃预变性5 min;94 ℃变性1 min,51~64 ℃退火1~2 min,72 ℃延伸1 min,35个循环;72 ℃再延伸10 min。
1.5 PCR 产物克隆、鉴定和序列测定
PCR 产物用胶回收试剂盒回收后,与pMD-18T 载体连接,转化,筛选重组质粒;重组质粒用PCR 鉴定,每个样品挑取3个阳性质粒,交由上海Invitrogen 公司进行序列测定。
1.6 序列拼接和分析
应用Lasergene V7.1 DNAstar 软件进行序列拼接和氨基酸序列推导,并用Mega 5.0 软件绘制gB、gC、gD和gE基因系统发育基因进化树。
2 结果与分析
2.1 gB 基因进化分析
2.1.1gB基因扩增 用gB基因引物进行PCR 扩增,扩增出大小约2.8 kb 的条带,与预期片段大小相符(图1)。
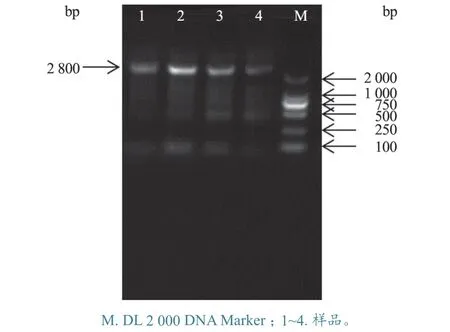
图1 PRV gB 基因扩增产物
2.1.2gB基因序列分析 测序结果显示,上海市2010—2015年分离毒株gB基因序列全长为2.8 kb,编码914个氨基酸残基组成的多肽。它们之间的核苷酸同源性和氨基酸同源性分别为99.6%~100% 和98.7%~100%,与GeneBank 上 国内外其他18 株的核苷酸同源性和氨基酸同源性分别为98.2%~100%和96.0%~100%(图2)。与国内Ea 毒株的氨基酸序列相比,存在6个位点的氨基酸一致性替换,即T82A、G393D、R451K、H560K、P735L、T737A。应用Mega 5.0 软 件中的Clustal W 方法,将测出的10 株PRVgB基因核苷酸序列与GenBank 上已登录的国内外其他18 株PRV 相应序列进行遗传距离分析,利用Neighbor-Joining 方法绘制了系统发育进化关系(图3)。基于gB基因进化树,PRV 毒株可划分为2个基因型Group1 和Group2。上海市2010年分离毒株与2012年以后分离毒株处于不同的进化分支,均与国内Ea 株亲缘关系较近,属于同一大进化分支(Group2),而与以欧美洲毒株为代表的Group1亲缘关系较远。
图2 PRV gB 基因核苷酸序列比较

图3 PRV gB 基因进化树分析结果
2.2 gC 基因进化分析
2.2.1gC基因扩增 用gC基因引物进行PCR 扩增,扩增出大小约1.4 kb 的条带,与预期片段大小相符(图4)。

图4 PRV gC 基因扩增产物
2.2.2gC基因序列分析 测序结果显示:上海市2010—2015年分离的PRVgC基因序列全长为1 464 bp,编码487个氨基酸残基,氨基酸序列具有典型的I 型囊膜糖蛋白特征。它们之间的核苷酸同源性和氨基酸同源性分别为94.9%~100%和99.2%~100%,与GeneBank 上国内外其他18 株的核苷酸同源性和氨基酸同源性分别为98.2%~100%和96.0%~100%。在对应Ea 株gC编码序列第186~211 位,上海市2010—2015年PRV 分离株与Ea 株多插入了21个核苷酸,并呈集中分布(图5)。应用Mega 5.0 软件中的Clustal W 方法,将测出的14 株PRVgC基因核苷酸序列与GenBank 上已登录的国内外其他72 株PRV 相应序列进行遗传距离分析,利用Neighbor-Joining 方法绘制了系统发育进化关系(图6)。基于gC基因进化树,PRV 毒株可划分为2个基因型Group1 和Group2。上海市2010年分离株与2012年以后分离株处于不同的进化分支,均与国内Ea 株亲缘关系较近,属于同一大进化分支(Group2),而与以欧美洲毒株为代表的Group1 亲缘关系较远。
2.3 gD 基因进化分析
2.3.1gD基因扩增 用gD基因引物进行PCR扩增,扩增出大小约1.5 kb 的条带,与预期片段大小相符(图7)。

图5 PRV gC 基因核苷酸序列比较
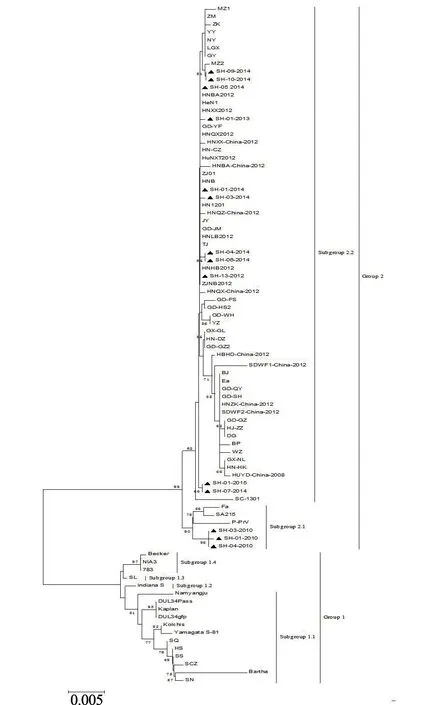
图6 PRV gC 基因进化树分析结果
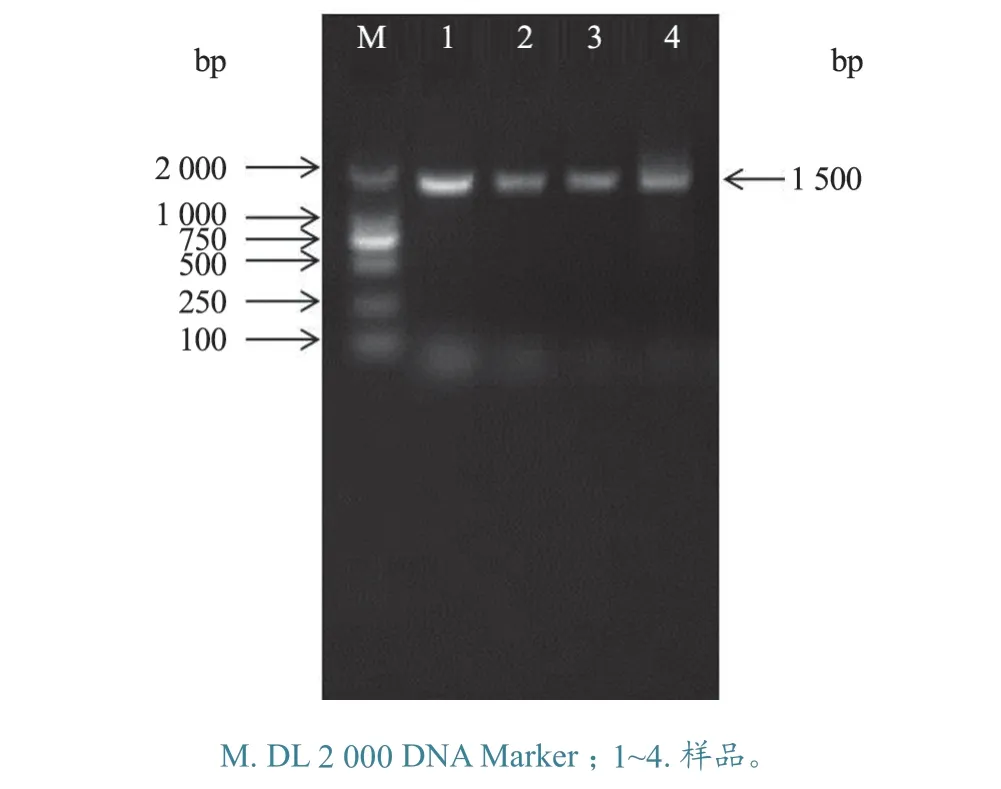
图7 PRV gD 基因扩增产物
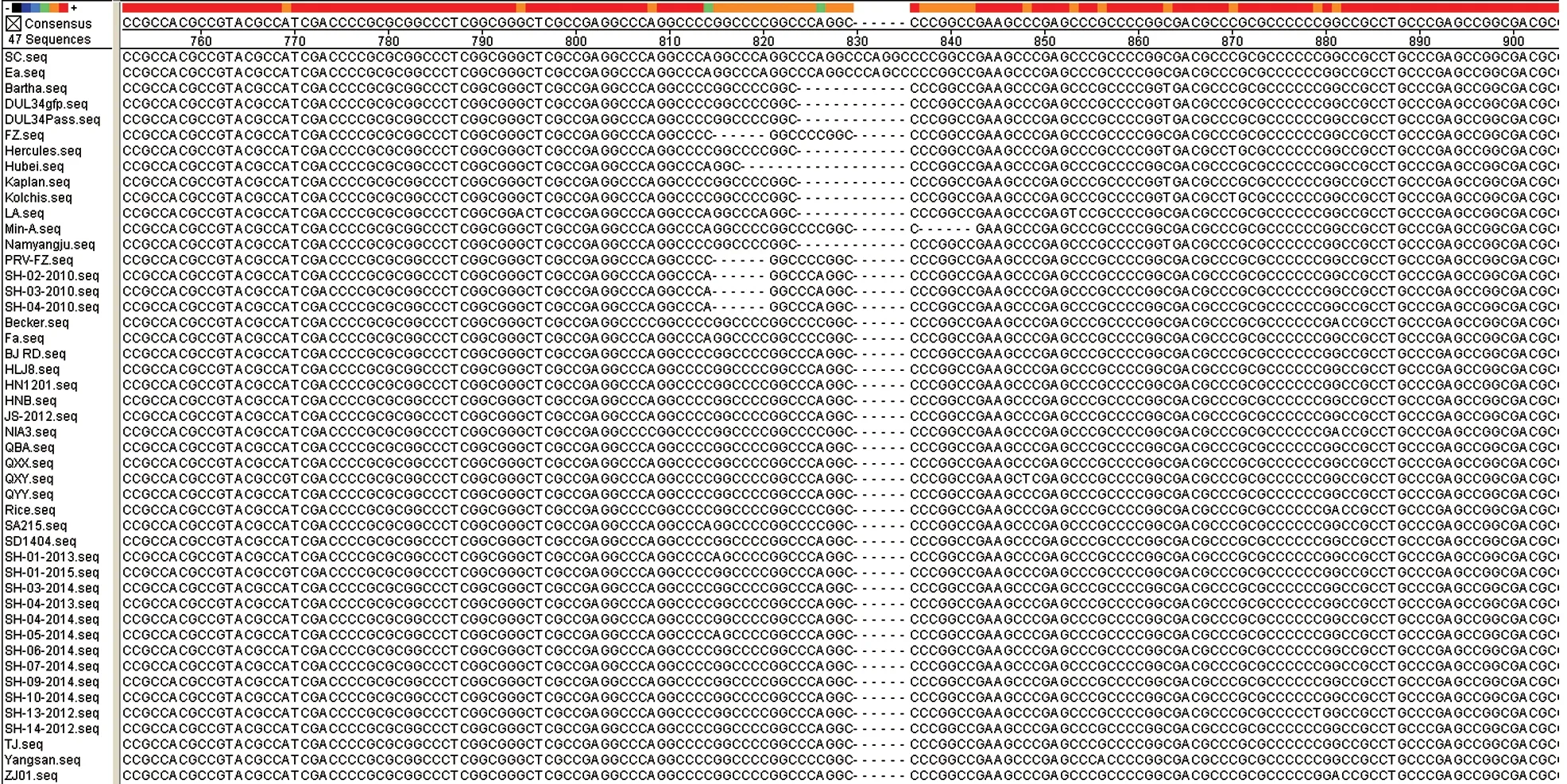
图8 PRV gD 基因核苷酸比对结果
2.3.2gD基因序列分析 测序结果(图8)显示:上海市2010年分离的3 株PRVgD基因序列全长为1 203 bp,编码400个氨基酸残基,而2012—2015年分离的12 株PRVgD基因序列全长为1 209 bp,编码402个氨基酸残基。相比,存在6个碱基(2个氨基酸)的缺失。这是由于gD基因802~837 nt处有1个C(A)GGCCC 重复高变区,对应的是267~279 位氨基酸Arg~Pro 的重复高变区。正是这一重复高变区的碱基缺失或插入,使得PRV-gD 的ORF 在1 194~1 215 nt 间变化,gD 前体的氨基酸残基为398~404个不等。2010—2015年分离的15 株PRV 的gD基因核苷酸及推导的氨基酸序列同源性分别为99.7%~100%和99.2%~100%;与其他32株PRVgD基因核苷酸及推导的氨基酸序列同源性分别为98.7%~100%和97.2%~100%。应用Mega 5.0 软件中的Clustal W 方法,将测出的15 株PRVgD基因核苷酸序列与GenBank 上已登录的国内外其他32 株PRV 相应序列进行遗传距离分析,利用Neighbor-Joining 方法绘制了系统发育进化关系。由图9 可知,PRV 毒株可以分为三大群(Group1、Group2 和Group3)。2010—2015年分离的15株PRV 与Min-A 株、Ea 株、FZ 株、SA215 株和LA 株亲缘关系最近,均属于Group3,而与以Bartha 株为代表的Group1 群和以Becker 为代表的Group2 群亲缘关系最远。

图9 PRV gD 基因进化树分析结果
2.4 gE 基因进化分析
2.4.1gE基因扩增 用gE基因引物进行PCR 扩增,扩增出大小约1.0 kb 的条带,与预期片段大小相符(图10)。
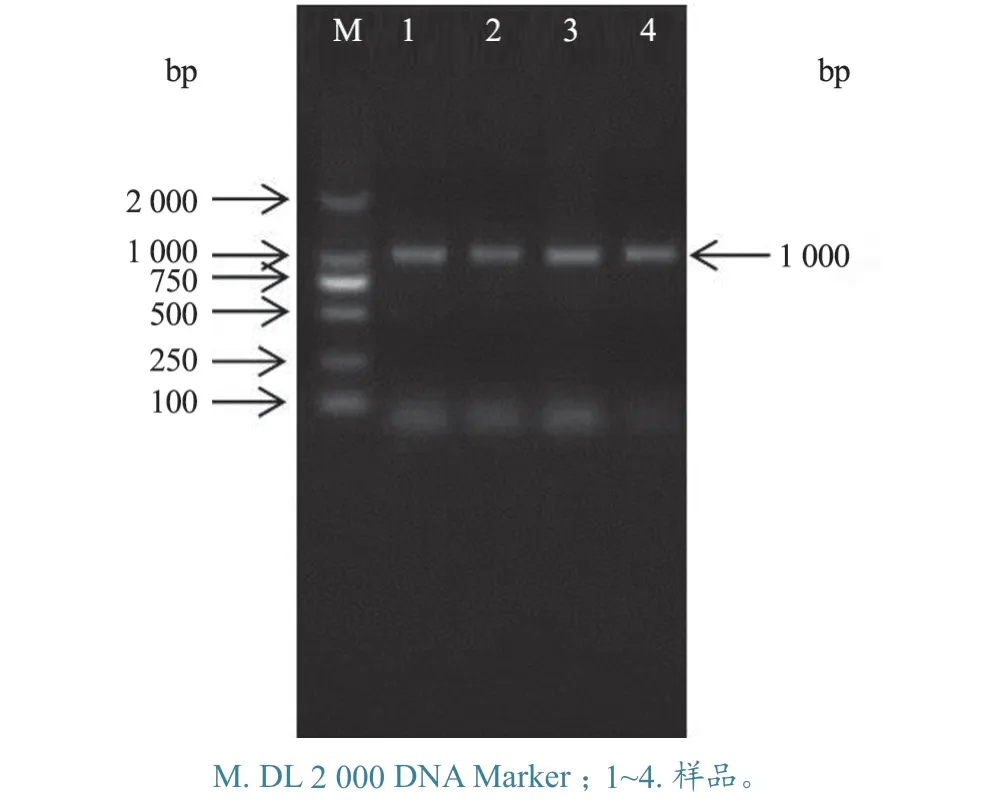
图10 PRV gE 基因扩增产物
2.4.2gE基因序列分析 用DNAstar V7.1 软件,将测出的26 株gE基因部分序列及推导的氨基酸序列与国内外39 株PRV 分离株序列进行同源性比较分析。结果显示:2010—2015年分离的26 株PRVgE基因核苷酸及推导氨基酸序列的同源性分别为99.5%~100%和99.0%~100%;与其他39 株PRV 的gE基因核苷酸及推导氨基酸序列的同源性分别为97.2%~100%和95.2%~100%,最低为NiA3 株。与国内Ea 毒株相比,2010—2015年分离的26 株PRV 存在多处碱基突变(图11)。应用Mega 5.0 软件中的Clustal W 方法,将测出的26 株PRVgE基因核苷酸序列与GenBank 上已登录的国内外其他39 株PRV 相应序列进行遗传距离分析,利用Neighbor-Joining 方法绘制了系统发育进化关系。由图12 可知,PRV 毒株可以分为两大群(Group1 和Group2),Group1 又分为两个亚群(Subgroup1.1 和Subgroup1.2),Group2 分为5个亚 群(Subgroup2.1、Subgroup2.2、Subgroup2.3、Subgroup2.4 和Subgroup2.5)。2010—2015年 分离的26 株PRV 均在Group2 中,其中2010年分离的4 株分布在Subgroup2.4 中,与Ea 株、P-PrV 株和Yangsan 株亲缘关系最近;2012—2015年分离的22 株均在Subgroup2.5 中,与国内2012年以后分离的流行毒株亲缘关系最近;均与以Min-A 株为代表的国内和东南亚分离株属同一大的进化分支(Group2),遗传关系相对较近;与国外欧美分离株Becker 株、Kaplan 株和NiA3 株分属不同的进化分支(Group1),亲缘关系较远。
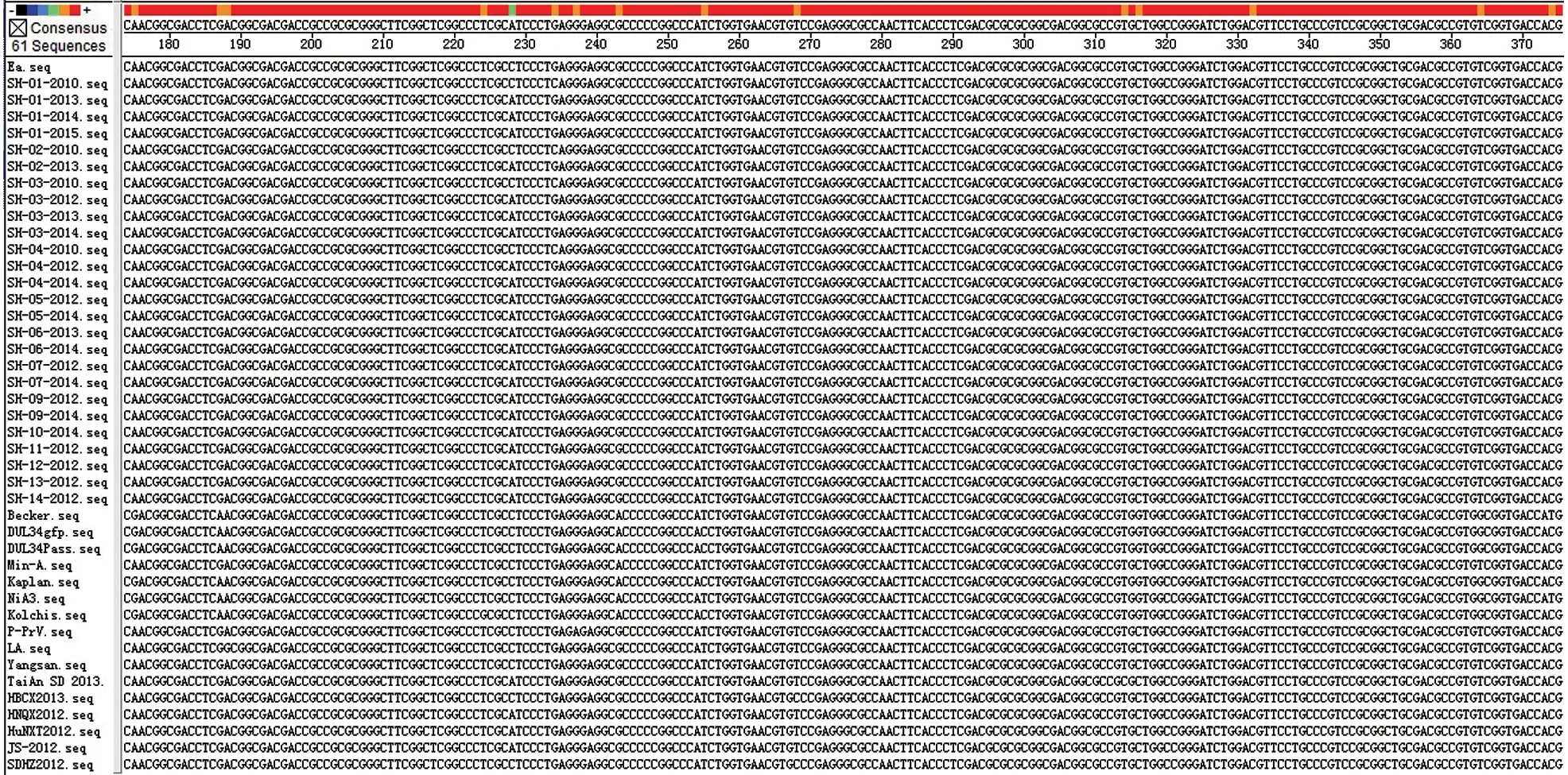
图11 PRV gE 基因核苷酸比对结果

图12 PRV gE 基因进化树分析结果
3 分析与讨论
3.1 PRV 相关毒力基因的PCR 扩增
PRV 基因组DNA 大小为150 kb,由于G+C含量高达73%以上,所以导致整个基因组结构复杂。局部DNA 复杂的二级结构经常影响PCR 的顺利扩增,同时对测序反应也提出了更高的要求。本研究开始使用宝生物LATaqE,结果未能很好地扩增出PRV 基因片段,随后提高退火温度,采用了宝生物PrimeSTAR HS DNA E。此酶具有高保真和高扩增效率,能直接高保真地扩增出高G+C 含量的PRV功能基因片段。建立的高保真扩增高G+C 含量基因的方法,对PRV 在构建重组病毒和克隆其余功能基因片段的研究方面提供了借鉴。此方法,也为扩增高G+C含量的基因片段提供一个不错的选择。
3.2 PRV 毒力相关基因分析
本研究对上海市2010—2015年分离的PRV 流行毒株毒力相关基因gB、gC、gD和gE进行了克隆和测序。测序分析显示,PRV 不同毒株间gB、gC、gD和gE基因在核苷酸和氨基酸水平的同源性均较高,但在高度保守的基础上,仍存在一些差异,部分差异具有特征性,如gB基因的集中突变区和gC编码序列186~211 bp 处集中多插入了21个核苷酸序列等。在疱疹病毒中,gB 蛋白属于最保守的糖蛋白之一。该糖蛋白不仅可诱导产生高滴度的中和抗体,而且对病毒的感染过程是必须的。PRV gB 蛋白的主要抗原表位已比较清楚。gB 蛋白分别在59~129、214~289、507~734 aa 处存在优势抗原位点[8];氨基酸变异位点主要集中在50~100氨基酸区段。本研究分析出的集中突变区正好位于此区域内。
PRVgD基因在803~837 bp 处有一重复高变区。在目前已发表的毒株gD基因序列中,只有Ea 株的gD基因最长,为1 215 bp,编码405个氨基酸的gD 前体;Rice 株与NiA3 株在这相应区域缺失6个核苷酸(AGCCCC),均为1 209 bp,编码402个氨基酸的gD 前体;LA 株与Kaplan株的这重复高变区均在相应区域缺失12个核苷酸(AGGCCCAGCCCC),均为1 203 bp,编码400个氨基酸的gD 前体;Hubei 株在这重复高变区有2 处缺失,在803~815 位缺失12个核苷酸(GGCCCAGCCCCA),在831~817 位缺失6个核苷酸(AGCCCC),共计缺失18个核苷酸,其gD基因长仅为1 197 bp,编码398个氨基酸的gD前体[9]。而上海市2010年分离的3 株PRVgD基因序列全长均为1 203 bp,编码400个氨基酸残基,而2012—2015年分离的12 株PRVgD基因序列全长均为1 209 bp,编码402个氨基酸残基。
gE基因在PRV 基因组中并不是最保守的基因。但PRV 本身遗传变异性不大,这不仅体现在PRV 只有1个血清型,而且也体现在基因水平上。gE基因的高同源性在一定程度上也体现了PRV 基因组的保守性。研究发现,碱基序列在1 037~1 407间的同源性高达100%,是gE基因中最保守的序列,编码的429~453 位氨基酸是疏水的膜锚定片段[10]。这个功能可能对于gE 蛋白非常重要。
3.3 PRV 毒力相关基因进化分析
基于gB、gC、gD和gE基因的进化树分析显示:欧美分离株可归为一大分支(Group 1),为基因I 型;上海市2010—2015年分离株,中国分离株与韩国分离株可归为另一个大支(Group 2),为基因II 型;2010年上海市分离毒株与以Min-A株和Ea 株为代表的毒株亲缘关系较近,处于同一小的分支;2012—2015年上海市分离毒株与2012年国内分离的流行毒株亲缘关系较近,与以Min-A株和Ea 株为代表的毒株亲缘关系较远,处于另一分支。由此可见,2012年后分离毒株的几个相关毒力基因均发生了变异,出现了PRV 变异株,从而导致了2013年伪狂犬病疫情的出现。
