南瓜银叶突变体48a的表型特征及其遗传学分析
2020-05-21汪端华杨建国李倩吴双花何长征马艳青
汪端华 杨建国 李倩 吴双花 何长征 马艳青


摘 要: 南瓜银叶突变体48a是在嫩食型中国南瓜中分离筛选到的稳定遗传自交系,银色叶不仅可以作为标记性状应用到育种中,还可为南瓜抗虫、抗病、耐寒等一系列研究提供重要的材料基础。笔者对银叶突变体的表型特征及叶片的解剖结构进行鉴定分析,发现植株整体长势、熟性与野生型无明显差异;成熟叶片正面全部呈银灰色,叶绿素含量明显降低,叶片上表皮细胞与栅栏细胞间明显剥离,存在明显的空隙。利用突变体48a和野生型49a南瓜自交系构建的六世代遗传群体,调查发现F2的绿叶与银叶符合3∶1的分离比,回交群体BC1P1分离比符合1∶1,表明南瓜银色叶性状由单隐性基因控制。
关键词:南瓜; 银叶突变体; 表型特征; 遗传学分析
中图分类号:S642.1 文献标志码:A 文章编号:1673-2871(2020)04-012-05
Abstract: Pumpkin silver leaf mutant 48a is a stable genetic inbred line isolated from tender pumpkin (Cucurbita moschata Duch.),which can be used not only as a marker trait in breeding, but also provide a series of important information such as insect resistance, disease resistance, cold tolerance. In this study, the phenotypic characteristics of silver leaf mutants and the anatomical structure of the leaves were identified, and we found that there was no significantly difference on growth and maturity between mutant 48a and wild type, but the content of chlorophyll was significant decreased and a clear gap exsisted between the upper epidermal cells and the fence cells in mutant 48a. From analysis of the sixth-generations group builded by mutant 48a and wild-type, the result shows that F2 green leaves and silver leaves was 3∶1 separation ratio, and 1∶1 separation ratio between the backcross population BC1P1, indicated that silver leaf traits were controlled by single recessive gene.
Key words: Pumpkin; Silver leaf mutant; Phenotypic characteristics; Genetic analysis
南瓜葉片银斑是南瓜中的常见性状,主要表现为叶脉的腋处带银白色斑纹,俗称“银斑叶”,是受单个显性基因控制的一种斑纹[1-3],区别于烟粉虱Bemisia tabaci B型(又名银叶粉虱B. argentifolii)诱发的葫芦科作物叶片正面全部呈银白色的银叶病[4-5]。2016年笔者在嫩食型中国南瓜资源中分离出1株完全银色叶突变体,自交后5代后形成可遗传的稳定银叶自交系,同时在同一材料中分离出稳定的普通绿叶自交系,田间种植观察发现银斑叶南瓜具有很好的避蚜和抗病毒病的效果,这为南瓜抗虫育种提供了新的思路。笔者对银叶突变体的表型特征、叶片解剖结构进行了鉴定分析,并构建了银叶六世代遗传群体进行遗传学分析,为后续的南瓜银色叶基因定位和避蚜上的应用奠定基础。
1 材料与方法
1.1 材料
在嫩食型中国南瓜材料的自交后代中,分离出具有完全银叶的突变体48a和普通绿叶野生型材料49a,经过5代自交,形成稳定的自交系。以突变体48a和野生型49a为亲本,构建六世代遗传群体,2018年4月将银叶南瓜亲本P1、绿叶南瓜亲本P2、F1、F2、BC1P1、BC1P2种子同时种于湖南省蔬菜研究所高桥试验田中。种子采用穴盘育苗,幼苗生长至3~4片真叶时定植,株行距50 cm×100 cm人字架栽培,进行统一的肥水管理,用于银叶和绿叶南瓜农艺学性状调查和银叶性状的遗传学分析。
1.2 方法
1.2.1 突变体48a的表型及农艺性状观察 突变体48a在苗期即可观察到明显的银叶表型,定植后到成株期都为银叶表型,6月中旬成株期,对主蔓长、主蔓粗、叶片长度、叶片宽度、叶柄长等生物学性状进行调查;同时在生育期内对其始花期(小区内50%植株开花的时期)、雌花数量进行调查,本试验分别调查测量了突变体和野生型南瓜材料各10株。
1.2.2 遗传学分析方法 定植后30 d左右,分别统计双亲、F1、F2、BC1P1、BC1P2等世代各群体银色叶和绿色叶的植株数量,然后通过卡平方测验,分析控制南瓜银叶性状基因的遗传特性。
1.2.3 叶绿素含量测定方法 待植株生长到成苗期,定植后30 d左右,分别取突变体48a和野生型49a的顶端嫩叶和相同位置的成熟叶片用于检测叶绿体色素含量。参考Arnon法[6],具体如下:随机称取叶片0.1 g,剪碎,装入15 mL带塞的玻璃试管中,加入10 mL的80%丙酮,黑暗浸泡24 h,至组织发白。将提取液在岛津公司的 UV-1800分光光度计上测定663、645、470 nm波长的OD值,计算Ca、Cb、Cx·c和CT(mg·L-1),公式如下:
Ca=12.21×OD663-2.81×OD645;
Cb=20.13×OD645-5.03×OD663 ;
Cx·c=(1 000×OD470-3.27×Ca-104×Cb)/229;
CT=Ca + Cb。
式中,Ca、Cb和CT分别表示叶绿素a、叶绿素b和总叶绿素浓度;Cx·c为类胡萝卜浓度,按下式计算组织中各色素含量。
叶绿体色素含量(mg·g-1)=(色素浓度×提取液体积×稀释倍数)/样品鲜质量。
1.2.4 叶片解剖学观察方法 定植后30 d左右,选取生长一致的突变体48a和野生型49a植株相同部位的成熟叶片,用于叶片解剖学观察。首先用FAA固定液固定,番红-固绿染色,RM2016病理切片机进行切片,切片放入干净的二甲苯透明5 min,中性树胶封片,NIKON ECLIPSE E100光学显微镜镜检,NIKON DS-U3成像系统进行图像采集分析。用测微尺测量叶片横截面的厚度、上下表皮厚度、栅栏组织厚度、海绵组织厚度,叶片结构紧密度(CTR)、疏松度(SR)的计算,参考田丽波等[7]的方法。
CTR/% =(栅栏组织厚度/叶片厚度)×100;
SR/%=(海绵组织厚度/叶片厚度)×100。
2 结果与分析
2.1 突变体48a的表型与农艺性状分析
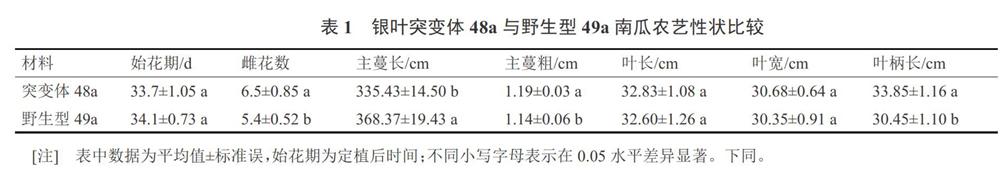
突变体48a和野生型49a进行表型对比观察。子叶期,银叶突变体和绿叶野生型子叶均表现为正常的绿色,子叶颜色无明显差别;但进入真叶期,突变体48a长出真叶为绿色,带明显银斑(图1-A),随着真叶逐渐长大,叶正面银灰色逐渐加深,植株长大成熟后,叶片正面全部为银灰色(图1-B、C、E),背面仍为绿色(图1-D)。
由成熟期的田间比较(图1-B)可以看出,除了叶色有明显区别外,突变体48a与野生型49a南瓜整体长势和熟性差别不大,对两者农艺性状测量数据显示,两者的始花期、叶长、叶宽等指标无明显差异。其中突变体48a南瓜主蔓长度(335.43±14.50)cm,比野生型49a低8.94%,而突变体48a主蔓粗度(1.19±0.03)cm,比野生型49a高4.39%,整體长势与野生型49a差别不大(表1)。
2.2 遗传学分析
构建六世代遗传群体,用目测分级的方法探究南瓜叶颜色的遗传规律,F1为绿叶、卡平方检验结果表明F2绿叶与银叶符合3∶1的分离比,回交群体BC1P1分离比符合1∶1,BC1P2植株叶颜色全为绿色, 南瓜银色叶性状符合1对显一隐性基因的分离规律,为完全隐性遗传(表2)。
2.3 叶绿素含量分析
由图2可以看出,突变体48a与野生型49a间叶绿素相比,叶片叶绿素含量明显减低,但嫩叶与成熟叶间的叶绿素无明显差别。从成熟叶来看,突变体48a叶片中,叶绿素和类胡萝卜素含量均显著低于野生型叶片,其中叶绿素含量的变化主要由于叶绿素a含量变化导致,叶绿素b含量相对于绿叶野生型叶片无明显差异。
2.4 突变体48a解剖学特征
从叶片横切面可见(图3-A)银叶突变体48a叶片的上表皮细胞与栅栏组织、下表皮细胞与海绵状组织之间有明显的空隙,其中上部空隙更大。从图3-B 可见绿叶野生型49a叶片的上表皮细胞与栅栏细胞,下表皮细胞与海绵状的叶肉细胞之间连接紧密,没有明显的空隙。
对突变体48a与野生型49a的叶片厚度、表皮细胞厚度、栅栏组织厚度、海绵组织厚度进行测量(表3),结果表明,突变体48a海绵组织厚度(44.2±3.00) μm,显著低于野生型49a,叶片厚度、上表皮厚度、下表皮厚度、栅栏组织厚度等指标两者间无显著性差异,通过测算,突变体48a叶片结构紧密度(CTR)32.06%,显著高于野生型49a,叶片叶片结构疏松度(SR)40.11%,显著低于野生型49a(表3)。
3 讨 论
突变体48a的银色叶可能是叶片结构变化所致,一般叶斑/片颜色变化产生原因主要分两类,色素的变化(叶绿素/花青素等)和叶片结构的变化(表皮细胞/空隙结构)。前人研究普遍认为银斑是叶片结构变化导致的,由于上表皮组织与栅栏组织之间的空隙,使光线达到绿色组织时候发生二次反射,在叶片上表皮形成多边形的光反射,而不是正常绿叶上皮细胞形成的白色点状光反射,使得叶片呈现偏灰白色,即银色的叶片/叶斑[8-10]。本研究结果表明,银叶突变体48a的叶绿素含量比野生型49a虽有显著降低,但也不会造成叶面显示银灰色,从突变体48a叶片解剖学特征显示,上表皮细胞与栅栏组织、下表皮细胞与海绵状组织之间有明显的空隙,其中上部空隙更大,这与张慧杰、张站备等[11-12]对西葫芦银斑研究一致,但他们的研究主要是基于西葫芦银叶病的叶片结构,突变体48a与西葫芦银叶病叶片在结构上虽然相似,但西葫芦银叶病会造成植株长势弱、植株矮小、生长点皱缩等特征[4],而突变体48a与野生型49a相比,除叶片呈银灰色外,植株整体长势正常,与野生型差别不大。所以突变体48a叶绿素含量明显减低,但长势正常的原因值得进一步研究。
银叶突变体48a的银叶表型为单基因的隐性遗传,与Lopez-Anido等[13]研究结果一致,他们在西洋南瓜和中国南瓜杂交后代中发现了1个完全银叶,研究表明由隐性基因grl控制。而Coyne[14]和Paris[15]等研究认为,南瓜银斑叶受M基因的显性遗传,且认为影响银叶性状变异的主要因素有3个,(1)细胞结构:V基因作用最强的部位是位于叶脉的腋处的细胞。(2)修饰基因:影响了M基因的时空表达。(3)包括温度/干旱在内的环境因素。据笔者田间试验表明,突变体48a的银叶性状,不受环境影响,在不同季节、不同环境下种植均显示完全银叶,也不受时空表达影响,在全生育期内均显示银色叶。目前南瓜的基因组测序已经完成[16],下一步拟利用全基因组混池测序的方法,进行银叶基因的快速定位,为后续研究该基因在南瓜中的作用机理奠定基础。
参考文献
[1] BHARATHAN N,NARAYANAN K R,MCMILLAN J T.Characteristics of sweet potato white fly-mediated silver leaf syndrome and associated double-stranded RNA in squash[J].Phytopathology,1992,82(2):136-141.
[2] SCOTT D H,RINER M E.A mottled leaf character in winter squash.[J].Heredity,1964,37:27-28.
[3] SCARCHUK J.Inheritance of light yellow corolla and leafy tendrils in gourd (Cucurbita pepo var.ovifera Alef)[J].1974,9:464.
[4] 張志轩,陈建芳,王雅丽,等.西葫芦银叶反应的原因及防治[J].长江蔬菜,2002(6):24.
[5] 胡敦孝,吴杏霞.银叶粉虱发生的指示植物西葫芦银叶[J].植物检疫,2001,15(3):132-136.
[6] 赵世杰,苍晶.植物生理学实验指导[M].北京:中国农业出版社,2015:77-80.
[7] 田丽波,商桑,杨衍,等.苦瓜叶片结构与白粉病抗性的关系[J].西北植物学报,2013,33(10):2010-2015.
[8] HORNER H T.Peperomia leaf cell wall interface between the multiple hypodermis and crystal-containing photosynthetic layer displays unusual pit fields[J].Annals of Botany,2012,109(7):1307-1316.
[9] SHIFRISS O.Reflected light spectra from silvery and nonsilvery leaves of Cucurbita pepo[J].Cucurbit Genetics Cooperative Report,1983,6:89-90 .
[10] ZHANG Y,HAYASHI T,HOSOKAWA M,et al.Metallic lustre and the optical mechanism generated from the leaf surface of Begonia rex Putz.[J].Scientia Horticulturae,2009,121(2):213-217.
[11] 张战备,张慧杰,段国琪,等.西葫芦叶上遗传性银斑与银叶病病斑光合生理及解剖特征比较[J].中国生态农业学报,2007,15(6):123-125.
[12] 张慧杰,段国琪,张战备,等.西葫芦银叶病叶片的光合生理与解剖学特征[J].植物病理学报,2005,35(4):327-332.
[13] LOPEZ-ANIDO F,COINTRY E,FIRPO I,et al.Inheritance of gray leaf color in a material derived from a Cucurbita maxima Duch.× C. moschata Duch. hybrid[J].Report-Cucurbit Genetics Cooperative,2002,25:46-48.
[14] COYNE D P.Inheritance of mottle-leaf in Cucurbita moschata Poir[J].Horticultural Science,1970(5):226-227.
[15] PARIS H S,HANAN A,BAUMKOLER F,et al.Assortment of five gene loci in Cucurbita pepo[M]//LEBEDA A,PARIS H S.Proceedings of Cucurbitaceae,Olomouc:Palacky University,Olomouc,Czech Republic,2004:389-394.
[16] SUN H,WU S,ZHANG G,et al.Karyotype stability and unbiased fractionation in the paleo-allotetraploid Cucurbita genomes[J].Molecular Plant,2017,10(10):1293-1306.
